Halliste luha taimkatte muutustest (0)
Esitatud küsimused
- Kui palju on Eestis puisniite?
Tartu Ülikool
Loodus- ja tehnoloogiateaduskond
Tartu Ülikooli Ökoloogia ja Maateaduste Instituut
Geograafia osakond
Magistritöö
HALLISTE LUHA TAIMKATTE MUUTUSTEST
Eha Puusild
Juhendajad: Prof. Tõnu Oja
MSc. Laimi Truus
Tartu 2008
SISUKORD
1. SISSEJUHATUS...................................................................................................................3
1.1. Pool-looduslike koosluste mõiste ja väärtus................................................................3
1.2. Luharohumaade mõiste, kujunemine ja looduslikud tingimused (taimekooslused
ja levik) ..................................................................................................................................5
1.2.1. Luhakooslusi mõjutavad tegurid ..............................................................................6
1.2.2. Rohumaade püsimist mõjutavad tegurid ..................................................................8
1.2.3. Luharohumaade levik ...............................................................................................9
1.3. Põllumajandusliku maakasutuse muutustest 20. sajandil ......................................11
1.4. Looduskaitse Soomaa rahvuspargis ..........................................................................12
1.5. Halliste luha geobotaaniline uuritus ..........................................................................13
1.6. Eesmärgid.....................................................................................................................15
2. MATERJAL JA METOODIKA .......................................................................................16
2.1. Uurimisala üldiseloomustus........................................................................................16
2.2. Materjal ja metoodika.................................................................................................19
2.2.1. Kaardianalüüsil kasutatud materjal ja metoodika...................................................19
2.2.2. Taimkatte uuringute materjal ja metoodika............................................................22
3. TULEMUSED.....................................................................................................................25
3.1. Niidu- ja metsakoosluste pindala muutused 20. sajandil.........................................25
3.2. Looduskaitseliste meetmete rakendamise mõju uurimisala luhakoosluste
pindalade levikule...............................................................................................................30
3.3. Uurimisala metsasus erinevatel maapinna kõrgustel...............................................33
3.4. Niiskustingimuste ja majandamisreziimi mõju luha taimkattele ...........................34
4. ARUTELU...........................................................................................................................38
4.1. Kaardianalüüsi tulemused..........................................................................................38
4.1.1. Halliste luha põllumajanduslikust maakasutusest ..................................................39
4.1.2. Looduskaitseliste meetmete rakendamise mõju uurimisala luhakoosluste pindalade
levikule. ............................................................................................................................40
4.2. Luhaniitude taimkatte uuringute tulemused ............................................................41
KOKKUVÕTE .......................................................................................................................42
SUMMARY.............................................................................................................................44
TÄNUAVALDUSED ..............................................................................................................46
KASUTATUD KIRJANDUS.................................................................................................47
LISAD
Lisa 1. ENSV Ministrite Nõukogu määrus 6. aprillist 1959. a. nr. 119
Lisa 2. Riikliku botaanilise keeluala "Halliste puisniit" kaart (1959)
Lisa 3. Taimkatte uuringualade paiknemine
Lisa 4. Truus, L., Puusild, E. 2008. Species richness, biomass production and recent
vegetation changes of Estonian floodplain grasslands. Käsikiri
2
1. SISSEJUHATUS
1.1. Pool-looduslike koosluste mõiste ja väärtus
Traditsiooniliselt majandatavate rohumaade kohta on kasutusel kolm enam-vähem võrdse
mahuga mõistet. Põllumajanduses ja maakatastri kõlvikuna on levinud väljend ,,looduslikud
rohumaad". See tähistab nende alade looduslähedust võrreldes kultuurrohumaadega.
Bioloogilises ja looduskaitselises erialakirjanduses on enam levinud terminid ,,pool-
looduslikud niidud" või ,,pool-looduslikud kooslused" mis arvestavad alade looduslikku
algupära ja püsimiseks vajalikku inimmõju. Viimastel aastatel on aga kasutusse tulnud (eriti
looduskaitselistes ringkondades) termin ,,pärandkooslused", mis viitab koosluse pikkadele
kasutamistraditsioonidele ja seotusele eestlaste rahvuskultuuriga. Samas ei väljenda see nende
alade looduslikku iseloomu ega ka seda, et tegu on rohumaadega (Luhamaa jt., 2001), seetõttu
eelistatatakse ja kasutatakse töös terminit ,,pool-looduslikud kooslused".
Pool-looduslikud kooslused etendavad väga olulist osa bioloogilise mitmekesisuse
säilitamisel. Siinkohal tuleks ära märkida nende kõrget liigirikkust, mistõttu neil on kõrge
teaduslik väärtus (Talvi, 2001).
Liigirikkus on kujunenud mitmetel erinevatel aja- ja ruumiskaaladel toimuvate protsesside
tagajärjel (Zobel, 1992; Grace, 1999). Väikesel skaalal on üheks oluliseks liigirikkuse ja
liigidünaamika mõjutajaks isenditevahelised interaktsioonid, mis sõltuvad võsude
horisontaalsest paigutusest ja need toimuvad lähimate naabrite vahel. Suuremaskaalalistest
teguritest on oluliseks koosluse liigiarvu mõjutajaks selle liigifond ehk nende liikide hulk, mis
on võimelised antud koosluses kasvama (Taylor jt., 1990; Zobel, 1992, 1997; Eriksson, 1993;
Pärtel jt. 1996; Pärtel & Zobel, 1999).
Luhaniidud on tekkinud lammimetsadest inimtegevuse tulemusena. Suuremate jõgede
(Emajõgi, Pärnu, Halliste, Pedja jõgi) ääres figureerisid need vähesel määral puisniitudena
juba atlantilisel kliimaperioodil. Laiemas ulatuses hakkasid aga lamminiidud kujunema alates
esimese aastatuhande keskpaigast e.m.a karjakasvatajate hõimude ilmumisel Eesti aladele
(Laasimer, 1965). Luhaniidud on suhteliselt vanad traditsioonilised kooslused, mistõttu on
neil suur liigifond, sest suurem hulk liike on pikema aja jooksul jõudnud seal kohaneda.
3
Kõrge liigirikkuse püsimisele on peale liigifondi hüpoteesi välja pakutud erinevaid selgitusi
(Grace, 1999; Barot, 2004), näiteks keskkonna heterogeensuse esinemine nii ajas kui ruumis
(Tilman, 1982), keskmine stressi, häirimise ja/või produktiivsuse tase neis kooslustes (Grime,
1973, 1979; Connell, 1978; Miller, 1982) ja teised mehhanismid, mis koos mõjutavad liikide
hulka.
Eestis on peamisteks pool-looduslike koosluste tüüpideks puisniidud ja -karjamaad,
lamminiidud ehk luhad, loopealsed ehk alvarid, rannaniidud, soo- ja soostunud niidud, palu-
ja nõmmeniidud ning lagedad aruniidud. Neli esimest on meil kõige tuntumad pool-
looduslikud kooslused, kuid ka ülejäänud on maastikulise ja liigilise mitmekesisuse
säilitamisel olulised (Luhamaa jt., 2001).
Pool-looduslikud kooslused on kadumas nii Eestis kui ka mujal maailmas, kuna neid ei peeta
majanduslikult olulisteks. Seetõttu on nende kaitse maailma praktilises looduskaitses
aktuaalne mitmel põhjusel. Üheks põhjuseks on eelpool nimetatud koosluste suur bioloogiline
mitmekesisus ja vajadus selle säilitamiseks. Teiseks oluliseks ajendiks on UNESCO maailma
kultuuri- ja looduspärandi kaitse konventsioon, mis kutsub üles hoidma maailma rahvastele
unikaalseid ja korvamatuid väärtusi. Viimaste hulka kuuluvad ka meie pool-looduslikud ehk
pärandkooslused, mis on looduse ja inimese ühislooming ning väärtuslikud nii teaduslikust,
kultuurilisest kui esteetilisest seisukohast. Soomaa rahvuspark on aastast 1998 UNESCO
maailmapärandi looduspärandi nimekirja nominandiks. Seega on meie ülesandeks kaitsta,
taastada ning hoida selliseid kooslusi tulevastele põlvedele. Pärandkoosluste traditsiooniline
majandamine on aga tänapäeval suurelt osalt võimalik vaid riigipoolse abiga. Oluline roll
nende säilitamisel on riiklikel looduskaitse asutustel, samuti erinevatel seltsidel ja ühingutel,
mille hulgast võiks esile tõsta Pärandkoosluste Kaitse Ühingut, mis loodi 1997. aastal.
4
1.2. Luharohumaade mõiste, kujunemine ja looduslikud tingimused (taimekooslused ja
levik)
Luha e. lammirohumaadeks nimetatakse jõgede orgudes (lammidel) paiknevaid, pikemat või
lühemat aega üleujutatavaid taimekooslusi. Luharohumaade all võib mõista ka järvede
üleujutatavaid kaldaalasid. Tavaliselt on viimastel setete hulk tähtsusetu ja seepärast
nimetatakse luha taimekooslusteks eeskätt neid, mis esinevad jõgede luhtadel (Laasimer,
1965).
Luharohumaadel võib kasvada ka üksikuid suuri puid ning põõsaid, enamasti vahetult jõgede
kallastel. Selliseid üksikute vanade puudega luhaniite nimetatakse ka lammipuisniitudeks,
kuid puistu hõreduse tõttu ei saa neist paljusid liigitada klassikaliste puisniitude hulka
(Luhamaa jt., 2001).
Lammimuldade kujunemise põhiliseks tingimuseks on jõe veetaseme kõikumistest tingitud
perioodiline üleujutus. Jõevesi kannab endaga kaasa mitmesuguse suurusega mineraalosakesi
(liiv, tolm, ibe), taimejäänuseid ja huumusaineid. Mida suurem on voolu kiirus, seda rohkem
ja suuremaid osakesi vesi endaga kaasa toob. Peale hõljumi sisaldab tulvavesi molekulaarselt
ja kolloidselt lahustunud aineid, milles on esikohal huumushapped. Need pärinevad tavaliselt
soodest ja annavad veele pruuni värvuse (Reintam, 1962).
Luhtadel eristatakse kolme ökoloogiliste tingimuste poolest erinevat vööndit, mille
kujunemine on seotud alluviaalse sette hulga ja üleujutuse ulatuse erinevustega: 1) sängiäärne
luht, kus moodustuvad kihilised lammimullad; 2) keskluht, kus kujunevad väga viljakad
teralised lammimullad; 3) terrassiäärne luht, kus settivad kõige peenemad osakesed ja setete
hulk on väike. Luhtade laiusest ja settelisuse intensiivsusest sõltub, kas seal esinevad kõik
eelpool nimetatud vööndid või mitte (Pork, 1959).
Et jõeorgude põhja tase on enamasti madal ja vähese kallakuga, siis esinevad valdaval osal
pindalast soostunud lammimullad ja vastavalt luhasoode taimekooslused. Kaldaäärsed
künnised on sageli kaetud pajustikega, mis soodustavad sette ladestumist ja on seetõttu
tüsedama huumuskihiga. Mullad on jämedamast sorteeritud materjalist, teralise struktuuriga,
mõnikord gleistunud, enamasti kõrge fosfori- ja lubjasisaldusega. Teiste luhavööndite
pindalaga võrreldes on aga nende osa väga väike. Peale kaldaäärsete künniste ehk
5
lammiaasade võib eristada lühiajaliselt, keskmiselt ja pikaajaliselt üleujutatavaid luhaniite,
mis vastaksid kõrge, keskmise ja madala taseme luhaniitudele. Lammi- ehk luhasood tekivad
jõesängist kaugemates osades, kus sette ladestumine on vaevalt märgatav, kuid vee äravool
reljeefi lameduse tõttu takistatud (Laasimer, 1965).
Laasimer (1965) on eraldanud neli luhaniitude ökoloogiliselt erinevat assotsiatsioonide
rühma: 1) kaldaäärsete künniste (lammiaasade); 2) lühiajaliselt kuni keskmiselt üleujutatud
luhaniitude; 3) pikaajaliselt üleujutatud luhaniitude ja 4) luhasoode assotsiatsioonide rühm.
Eesti tingimustes moodustavad luhaniitudest pindalalt suurema osa keskmiselt kuni
pikaajaliselt üleujutatavad niidud, mis üleujutuste tõttu on soostunud või muutunud
sooniitudeks.
1.2.1. Luhakooslusi mõjutavad tegurid
Porgi (1959) poolt läbi viidud uuringute kohaselt võime luhtadel eristada kolme suurvee faasi:
1) üleujutuse tulvafaas; 2) üleujutuse kõrgfaas; 3) üleujutuse alanemisfaas. Faaside kestus on
erinevatel jõgedel erinev.
Alluviaalse sette ladestumise tagajärjel muutub luha reljeef ning jõesäng asetub pidevalt
ümber. Jõe põrkeveeru poolsel küljel lähenevad luhad pidevalt jõele, kuid jõe liuveerul
ladestuvad settekihid ja need alad kerkivad ning eemalduvad jõest. Seetõttu läbivad kooslused
jõe kaldal mitmeid arengustaadiume. Järsematel nõlvadel jääb osa vahepealseid staadiume
ära, nende suktsessiooni rida on järgmine: Carex gracilis`e ass. kaldaäärne
variantDigraphis arundinacea (Phalaris arundinacea) ass. Carex gracilis`e
var.Ranunculus repens-Poa trivialis`e ass. või taimkatteta ribaDeschampsia caespitosa-
Festuca rubra ass. (Pork, 1964).
Kaldalähedastel aladel asenduvad taimekooslused teistega vastavalt niiskustingimuste
muutumisele. Jõesängi ümberasetumisega seotud ökoloogiliste tingimuste muutumiste suurt
osatähtsust näitab ka erineva lõimisega kihtidega kaetud sette- ning turbahorisontide sage
esinemine mullaprofiilis. Erinevate kihtide moodustumise ajal on valitsenud erinevad
ökoloogilised tingimused, mis mõjutasid suuremal või vähemal määral sealset taimkatet
(Pork, 1964).
6
Ressursikonkurents taimede vahel toimub toitainete ja valguse pärast. Suure liigitihedusega
niitudel peavad seal kasvavad liigid kuidagi vältima konkurentset väljatõrjumist. Häirimine
toimub liigirikastes kooslustes peamiselt regulaarse niitmise või karjatamise läbi. See
võimaldab edukalt konkureerida ka muidu konkurentselt nõrgematel liikidel, eeskätt
võsudevahelise asümmeetrilise valguskonkurentsi vähendamise kaudu (Grubb, 1986). Samuti
on regulaarselt niidetavates kooslustes võsud väiksemad ja seega kasvavad tihedamalt
(Diemer & Pfadenhauer, 1987; van der Maarel, 1988). Mida suurem on aga võsude arv, seda
suurem saab olla liikide arv väikesel skaalal (Abrams, 1995; Oksanen, 1996; Zobel & Liira,
1997; Pärtel & Zobel, 1999; Stevens & Carson, 1999).
Karjatamine algas luhtadel varem kui niitmine, tõenäoliselt juba 3000-4000 aastat tagasi.
Luhatingimustes on karjatamise mehhaaniline mõju väga tugev. Karjatamise suhtes tundlikud
taimeliigid hävivad, tolerantsed liigid aga jäävad püsima. Niitmise ja karjatamise tulemusena
suureneb luhtade taimkattes kõrreliste ja tarnade osatähtsus (Pork, 1964).
Raskete heinakoristusmasinate mõju on toimelt sarnane karjatamisele. Eriti tugevalt
mõjutavad masinad märjemaid luhaalasid. Traktori rattajälgedes on märgata tarnade ja
väärtuslike heintaimede vähenemist. Seal levivad seemneliselt paljunevad lehtrohud. Rohustu
hõrenemine, madal kasv ja liigilise koosseisu muutus on märgatav veel 3-4 aastat peale
traktori sõitmist. Kuivematel luhtadel on masinate mõju väiksem kuid paljudel neist on
traktorijäljed märgatavad veel mitmelgi aastal. Eriti tugev on masinate mõju heinaveoteede ja
luhtade pealesõiduteede ääres (Pork, 1981).
Luhtade taimkattele avaldab mõju ka jõe veereziimi muutmine: kas jõe süvendamine või
veetaseme tõus. Jõgede süvendamise tulemusena alaneb ümbritsevate alade põhjavee tase
ning selle tagajärjel niitude üldine niiskusaste. Põhjavee taseme alanemise kõrval alaneb ka
üleujutus ja väheneb alluviaalse sette hulk. Sellest tingituna muutuvad keskkonnatingimused
pärast jõe süvendamist oligotroofsemaks ja eutroofsed taimekooslused asenduvad
oligotroofsematega. Pikaajaliselt üleujutatud aladel, kus levisid kõrgekasvulised tarnad,
asenduvad need madalakasvuliste ja hõredapuhmikuliste tarnadega. Tarnakooslused
asenduvad kõrreliste kooslustega. Eriti tugevalt avaldub kuivenduse mõju luhasoodes, kus
Carex elata assotsiatsioon asendub hästi arenenud samblarindega Carex lasiocapra-
Drepanocladus revolvens`i assotsiatsiooniga (Pork, 1964). Jõe veetaseme tõus põhjustab
põhjavee üldise taseme tõusu ning üleujutuse kestuse suurenemist. Luhtade taimekooslused
7
muutuvad märjemaks, mistõttu kõrreliste kooslused asenduvad tarnakooslustega ja settelistele
aladele iseloomulike kooslustega (Pork, 1964).
1.2.2. Rohumaade püsimist mõjutavad tegurid
Mitmeaastased heintaimed loovad oma võsude reeglipärase vegetatiivse uuenemisega erilise
biotsönootilise horisondi rohustu. Niidutaimede maapealsete osade eluiga on tavaliselt
aasta, ainult mõnede liikide roomavad võsundid elavad kauem. Kuid nende juured ja risoomid
elavad 2-3 aastat või kauem, mille vältel osa neist sureb või uueneb. Seetõttu toimub maa-
aluste organite (risoomid, narmasjuured) kuhjumine mulla kõige ülemisse horisonti
kamardumine. Metsakooslustes on kamardumine nõrgalt arenenud. Heintaimede juurte
põhiline mass asub ülemises 0-20 cm tüseduses mullahorisondis ja ületab taimede maapealse
massi mitu korda (Krall, 1979).
Rohukamara tiheduse suurenedes kasvab ka juurte arv ja mass. Rööbiti orgaanilise aine
moodustumisega kamaras toimub seal ka taimejäänuste intensiivne lagunemine, lämmastiku
sidumine ja huumuse moodustamine, mille tagajärjel tõuseb mulla üldine viljakus.
Rohukamar on elupaigaks heintaimedega kohastunud organismidele: seentele, bakteritele,
mullafaunale. Kui ühtede taimeliikide produktsioon langeb, siis võib kaasneda teiste
taimeliikide produktsiooni tõus. Sümbiotroofidest on kõige tähtsamad mükoriisat
moodustavad seened. Enamik niidutaimi on mükotroofsed (Krall, 1979).
Niidurohustu püsimist soodustab tugev kamardumine, mis takistab osaliselt puude (ja ka teiste
taimede) seemnelist uuenemist. Lopsakas rohustu kasv takistab puutõusmete kasvu. Noored
puutaimed, mis on suutnud rohustust üle kasvada, hakkavad aga kiiresti sirguma ja võidavad
rohttaimi valguskonkurentsis. Selle tulemusena hakkavad rohumaad võsastuma ja metsastuma
(Krall, 1979).
Kuivematel luhaosadel levivad niitmise katkemise järel kiiresti pajupõõsad, eriti mustjas ja
kõrvpaju (Salix mysinifolia, S. aurita). Karjamaadena kasutavatel luhtadel levib aga hundipaju
(S. rosmarinifolia). Aeglasemalt levib luhal raudremmelgas (S. pentandra), sookask (Betula
pubescens), hall- ja sanglepp (Alnus incana, A. glutinosa) (Pork, 1959).
8
Märgatavad muutused toimuvad niitmise katkemise tagajärjel ka rohurindes. Muutub
rohttaimede liigiline koosseis, mis on tingitud kärpimise kui paljude liikide arengut allasuruva
faktori puudumisest. Paraneb varustatus toiteelementidega. Toiteelemendid, mis niidetavatelt
aladelt saagiga ära viidi, muutuvad varise lagunemise järel taimedele uuesti kasutatavaks.
Seetõttu on mitteniidetaval alal rohustu kõrgem ja lopsakam, selles kasvavad
lämmastikunõudlikumad liigid. Märgades kasvukohtades suureneb tunduvalt pilliroo
(Phragmites australis) ohtrus, kuna ta talub niitmist halvasti ja regulaarselt niidetavatel aladel
langeb sageli kooslustest välja. Niitmise katkemise järel hakkab pilliroog uuesti vegetatiivselt
arenema, muutudes aja jooksul dominandiks (Pork, 1981).
Mitteniidetavatel aladel hakkavad rohkem levima lehtrohud: soopihl (Potentilla palustris),
angervaks (Filipendula ulmaria), ubaleht (Menyanthes trifoliata), harilik metsvits
(Lysimachia vulgaris), kollane võhumõõk (Iris pseudacorus), suur tulikas (Ranunculus
lingua) jt. Ilmuvad ka mitmed niidetavatel aladel harva esinevad liigid: soo-nõianõges
(Stachys palustris), harilik maavits (Solanum dulcamara), ussilill (Naumburgia thyrsiflora),
mürkputk (Cicuta virosa), männasmünt (Mentha verticillata) jt. (Pork, 1981). Soomaa
luhtadel laiuvad valdavalt angervaksa- ja tarnaväljad.
1.2.3. Luharohumaade levik
Euroopas on luharohumaad levinud suuremate jõgede lammidel. Põllumajanduses viimase
sajandi jooksul toimunud muutuste tõttu on aktuaalsed nende majandamise ja taastamise
probleemid (Benstead jt., 1999).
Eestis võib luharohumaid sagedamini leida Kesk- ja Lõuna-Eestist, suurimad neist asuvad
Emajõe ja Pedja ning Koiva ja Mustjõe ääres. Lääne-Eesti ulatuslikumad luhad asuvad Kasari
jõgikonnas (Pork, 1984). Luharohumaade pindala on Eestis olnud suhteliselt väike. Laasimeri
(1965) andmetel oli 1960ndatel Eestis ca 83000 ha luharohumaid, mis moodustasid vabariigi
pindalast 1,8%. Suuremad neist asusid Tartumaal hõlmates 10,5% luhtade üldpindalast ehk ca
26200 ha. Suuremal pindalal esines luharohumaid veel Põlvamaa idaosas (5000 ha),
Valgamaal (5600 ha), Jõgevamaal (6300 ha) jne. Luharohumaade osatähtsus oli minimaalne
Hiiumaal, Saaremaal, Harjumaal ja Ida-Virumaal ning Pärnumaa põhjaosas (Laasimer, 1965).
9
1978-1981.a. inventuuri tulemusena hinnati luharohumaade üldpindalaks 27584 ha.
1960ndate andmetega võrreldes oli nende pindala vähenenud ligikaudu kolmandiku võrra.
Enim oli säilinud luharohumaid Läänemaal (4186 ha), Tartumaal (3935 ha),Viljandimaal
(3481 ha), Võrumaal (3043 ha), Valgamaal (2776 ha) ja Põlvamaal (2263 ha) (Leibak, Lutsar,
1996).
Aastail 1993-1996 inventeeriti projekti WETSTONIA käigus olulisemat looduskaitselist
väärtust omavaid luharohumaid. Kõrge ja keskmise looduskaitselise väärtusega
luharohumaade pindalaks hinnati ca 12500 ha. Maakondade kaupa jagunesid need järgmiselt:
Läänemaa 4550 ha, Tartumaa 2200 ha, Ida-Virumaa 1300 ha, Viljandimaa 1250 ha,
Põlvamaa 1000 ha, Valgamaa 750 ha, Võrumaa 700 ha, Pärnumaa, Jõgevamaa ja
Harjumaa - 200 ha ja Lääne-Virumaal 100 ha. Teistes maakondades olulise looduskaitselise
väärtusega luharohumaid ei registreeritud (Leibak, Lutsar, 1996).
1999-2000 tehtud inventuuride käigus hinnati lammirohumaade kogupindalaks 10 777 hekarit
(868 niiduala), millest kõrge ja keskmise väärtusega luhti oli kokku 8203 hektaril. Kuna
inventuur ei hõlmanud suuremaid kaitsealuseid luhti Soomaal, Alam-Pedjal ja osaliselt ka
Matsalus, on luhaniitude pindala ilmselgelt suurem. Paremini säilinud lamminiitude pindala
on hinnangute järgi 15000-20000 ha (Kukk, 2004).
10
1.3. Põllumajandusliku maakasutuse muutustest 20. sajandil
20. sajandi esimesel poolel erines põllumajanduslik maakasutus oluliselt tänapäevasest.
Tasasema reljeefiga ning kõrgema mullaboniteediga alad olid sajandite vältel suurelt osalt
põllumaadena kasutusel. Pool-looduslikud kooslused etendasid olulist rolli kariloomade
söödavarumisbaasina. Karjamaaks jäid sageli metsatukad, mis ei sobinud põllumaaks. Jõgede
kaldaalade luhad sobisid üleujutuste ja liigniiskuse tõttu vaid heinavarumiseks (Talvi, 2001).
Pool-looduslike koosluste majandamisega seotud traditsioone on lähemalt kirjeldanud O.
Loorits (2000; 2001).
Enne 1919. a. kuulus 42% maast ostutaludele ja 58% mõisatele. Mõisate maast oli ligi
veerand antud renditaludele. Seega oli väikemajapidamiste (74663 ostu- ja renditalu)
kasutuses 55,3% ja suurmajapidamiste (mõisate) valduses 44,7% üldisest maafondist.
(Kasepalu, 1991).
1940. a. maareformiga kuulutati maa riigi omanduseks, mis pidi tähendama maa muutumist
rahva omandiks. Tegelikult sai see maa ja vara sunniviisilise võõrandamise aluseks. 30
hektarit ületav maa tuli taludelt ära lõigata ja arvata riiklikku maatagavarasse uute talude
tegemiseks ning väiksematele juurdelõigete andmiseks. Talumaade vähendamisega kaasnes
kohati ka kariloomade äravõtmine ning põllutöömasinate tasuta võõrandamine. Kui väideti, et
nõukogude reform vähendas talude vahel valitsevat ebavõrdsust, siis tegelikult see nii ei
olnud. Maa ja inventari äravõtmine nõrgendas suuremate talude tootmisvõimet. See ei
aidanud väiketalupidajaid ja maata inimesi paremale järjele. Eesti talukultuuri hävitamine
viidi lõpule kollektiviseerimisega (Kasepalu, 1991).
1950ndatel läbi viidud kollektiviseerimine oli üks esimene puisniitude kadumise põhjus.
Peamine puisniitude pindala vähenemise põhjus oli suurtootmisele üleminekuga kaasnenud
käsitsitööst loobumine. Intensiivsemale põllumajandusele üleminek on olnud puisniitude
kadumise peapõhjus ka Lääne-Euroopas (Kukk, 2004).
Eestis aset leidnud maakasutuse ja maastiku mitmekesisuse muutustest on põhiliseks olnud
põllumajandusliku maa osatähtsuse vähenemine 65%lt 1918. aastal 30%ni 1994. aastaks.
Metsamaa osatähtsus on kasvanud vastavalt 21%lt 43%ni samas ajavahemikus (Mander,
Palang, 1998).
11
Põhilisteks maakasutust mõjutavateks jõududeks on peale looduslike protsesside olnud veel
maareformid, küüditamised, linnastumine ja põllumajanduse kontsentratsioon nõukogude
perioodil (Mander jt, 1994).
Viimase aastasaja jooksul on paljud niidud suurelt jaolt kinni kasvanud, kuna
traditsioonilistest maakasutusviisidest karjatamisest ja niitmisest on intensiivsemate
põllumajandusmeetodite kasuks loobutud. Järsud muutused traditsioonilises
põllumajanduslikus maakasutuses on seadnud ohtu aladele iseloomulikud taime- ja
loomaliigid (Hallanaro jt, 2002). Kunagistest laialt levinud pool-looduslikest kooslustest on
Euroopas järele jäänud vaid üksikud fragmendid (van Dijk, 1991).
1.4. Looduskaitse Soomaa rahvuspargis
Esimest korda rakendati looduskaitse meetmeid praeguse Soomaa rahvuspargi territooriumil
1959. aastal, mil ENSV Ministrite Nõukogu määruse nr. 119 (lisa 1) alusel loodi riiklik
botaaniline keeluala "Halliste puisniit" (lisa 2) ja määrati kindlaks selle piirid. Ühtlasi
kinnitati ka keeluala kaitse korraldamise eeskiri, mille kohaselt oli nimetatud keeluala
ülesandeks Halliste jõe luhal kasvava omapärase tamme-pärna-saare puisniidu säilitamine ja
sealsete taimekoosluste edasise arengu jälgimise võimaldamine. Keelatud oli puude raiumine
(välja arvatud sanitaarraied), karjatamine, taimede kogumine, ala ülesharimine või selle
üldilme muutmine ning jahipidamine kogu aasta vältel (välja arvatud röövloomade ja lindude
hävitamine organiseeritud korras kooskõlas jahipidamise eeskirjadega) (Kumari, 1960).
1993. a. 8. detsembril moodustati Eesti Vabariigi valitsuse määrusega nr. 387 Kikepera,
Kuresoo, Valgeraba ja Öördi sookaitsealade ning Halliste puisniidu botaanilise keeluala baasil
Soomaa rahvuspark. Selle ülesandeks on Vahe-Eesti edelaosa soode, lamminiitude ja metsade
kaitse (Allilender, 2000).
Soomaa rahvuspargi kaitse-eeskirja (2005) järgi kuulub Halliste luht rahvuspargi 29
sihtkaitsevööndi hulka, mis on loodud väljakujunenud või kujunevate looduslike ja
poollooduslike koosluste säilitamiseks.
12
Rahvuspargi algusaastatel tehtud niitude inventuuride käigus selgus, et ligikaudu 75%
Halliste luhaaladest on võsastunud. Ennustati jõe vasakkalda niidukoosluste täielikku
kadumist lähima aastakümne jooksul. Rahvuspargi loomise ajaks oli Halliste jõe
luhaniidukooslusest suhteliselt hästi säilinud vaid ca 10%. Leiti, et Halliste luha kõige
esinduslikemate piirkondade - Läti, Tõramaa ja Halliste puisniidu säilitamiseks tuleks
regulaarselt hooldada 130 ha suurust ala (Suurkask, 1996).
Soomaa rahvuspargis inventeeriti aastail 1996-1999 kokku ca 2000 ha luhaniite, mis
jagunesid 20 erineva niiduala vahel. Peaaegu 50% luhtade pindalast jäi Halliste jõe kallastele.
Väiksemad niidualad olid ca 25 ha, suuremad üle 200 ha suurused. Suurimad luhad olid
Sandra luht (245 ha) Raudna jõe kaldal, Vodi luht (236 ha), Tipu luht (199 ha) ja Lennuväli
(194 ha) Halliste jõe kaldal (Suurkask, 1999).
Luhad jaotati nelja majandamisklassi. Esimese, teise ja kolmanda majandamisklassi niite
plaaniti võimaluste piires hooldada. Neljanda majandamisklassi niitude hooldamist aga ei
peetud võimalikuks ning need alad olid määratud võsastuma. 1, 2. ja 3. klassi luhtade
kogupindalaks rahvuspargis hinnati ca 890 ha, mis moodustasid kokku 44% rahvuspargi
luhtade pindalast. 4. majandamisklassi luhti oli 805 ha. Kõige rohkem hooldamist vajav
territoorium jäi Halliste jõe kallastele kokku 471 hektaril, sellele järgnesid Raudna jõe luhad
206 hektariga, Tõramaa jõe luhad 115 hektariga ja Lemmjõe luhad 98 hektariga. Kokku peeti
soovitatavaks igaaastaselt hooldada ca 500 ha luhti, kuid seda teostati ca 2/5 minimaalsest
vajalikust territooriumist (Suurkask, 1999).
1.5. Halliste luha geobotaaniline uuritus
Esimese geobotaanilise ülevaate luha taimkatte kohta andis T. Lippmaa (1932) Edela-Eesti
loodust kirjeldavas töös "Beiträge zur Kenntnis der Flora und Vegetation Südwest-Estland".
Teine põhjalikum uurimistöö käsitletava ala kohta valmis H. Miku (1957) diplomitööna
"Halliste jõe luhtade taimkate (Tipu ja Riisa küla vahelisel alal)", milles anti ülevaade Halliste
jõe luhal esinenud taimekoosluste levikust, floristilisest koosseisust ja majanduslikust
kasutamisest. Põhilise osa tööst moodustas materjal, mis koguti Halliste jõe paremkaldal
esinevatelt luhaniitudelt. Kokku registreeriti 187 kõrgemat soontaimeliiki ja 18 samblaliiki.
Kogu kirjeldataval alal eraldati 20 taimekoosluse tüüpi. Kõige suurema levikuga märgiti
Deschampsia caespitosa (luht-kastevarre)-Carex panicea (hirsstarna), Carex panicea
13
(hirsstarna)-Carex goodenowii (C. nigra ssp. nigra, hariliku tarna alamliigi), Carex disticha
(lünktarna), Carex gracilis (saleda tarna) kooslusi.
Ülevaatlikke andmeid Halliste luha kohta kajastab ka T. Pullissaare diplomitöö "Materjale
Kasari, Halliste ja Navesti luhaniitude taimkattest" (1959). Välitööde käigus tehti 290
geobotaanilist analüüsi, neist Soomaa luhtadelt 85. Anti ülevaade luhal levivatest
taimekooslusest. Enamlevinud kooslusteks kõigil luhtadel olid tarnakooslused: Carex elata
(luhttarna), Carex disticha ja Carex gracilis kooslused. Kõrreliste kooslustest levisid Halliste
luhal Deschampsia caespitosa (luht-kastevarre)-Festuca pratensis (hariliku aruheina),
Sesleria caerulea (hariliku lubika) kooslused.
Soomaa luhtade uurimisega on põhjalikumalt tegelenud M. Suurkask. 1996. a. suvel
inventeeris ta Halliste jõe luhta ning puisniite ja andis ülevaate luhaala tähtsamate biotoopide
levikust, nende seisundist ja looduskaitselisest väärtusest. Kokku leidis M. Suurkask Halliste
luhalt 222 liiki soontaimi, sealhulgas 7 kolmanda kategooria kaitstavat liiki, Läti puisniidult
167 liiki ja Halliste puisniidult 171 liiki (Suurkask, 1996).
Soomaa rahvuspargi kaitsekorralduskava ettevalmistava projekti "Soomaa rahvuspargi
luhtade bioloogiline mitmekesisus ja majandamine" raames jälgiti nelja jõe Halliste,
Raudna, Tõramaa ja Lemmjõe luhtade seisundit, hinnati nende looduskaitselist väärtust ja anti
soovitusi edasiseks majandamiseks. Samasisuline on ka magistritöö "Soomaa rahvuspargi
lamminiitude taimkate ja suktsessiooniline seisund" (Suurkask, 1999).
Soomaa rahvuspargi Karuskose ja Lemmjõe lammimetsa taimekoosluste ja muldade toitainete
sisalduse dünaamikat erineva niiskusreziimiga aastatel on uurinud M. Karu (2001).
14
1.6. Eesmärgid
Käesoleva töö eesmärgid on:
1) anda luha kui Soomaa ühe võtmeökosüsteemi iseloomustus;
2) anda ülevaade luhakoosluste hüdroloogilistest ja troofsustingimustest ning nende seosest
majandustegevusega;
3) kaardimaterjali analüüsi rakendades uurida, kuidas on looduskaitse meetmed ning
muutused põllumajanduslikus maakasutuses mõjutanud uuritava piirkonna luhaniitude
pindala;
4) selgitada välja, millised on majandamise katkemise järel taimkattes toimunud muutused;
5) leida seoseid luhataimkatte struktuuri, maapealse biomassi, liigirikkuse ja eluvormide
jaotumise vahel erinevates niiskustingimustesja majandamisreziimidel.
15
2. MATERJAL JA METOODIKA
2.1. Uurimisala üldiseloomustus
Soomaa rahvuspark paikneb Madal- ja Kõrg-Eesti piiril Sakala kõrgustiku läänenõlval Pärnu
madaliku idaosas. Rahvuspargi pindala on 39 639 ha (Eesti Looduse Infosüsteem, seisuga
nov. 2005).
Kliima tingimustelt on Soomaa üleminekupiirkonnaks otseselt mere poolt mõjutatavalt alalt
(Läänemereline kliimavaldkond) Lõuna-Eesti tasandikulisele alale (Sise-Eesti
kliimavaldkond). Aasta keskmine õhutemperatuur on +4,5º kuni +5ºC, juulis +16,6º ja
veebruaris 6,6ºC. Sademeid langeb keskmiselt 670 mm aastas (Allilender, 2000).
Pinnamood selles piirkonnas on tasane, absoluutkõrgus on valdavalt 20-35 m ü.m.p. Tasane
reljeef ei soodusta pinnavee äravoolu, põhjustades soostumist ja jõgede üleujutusi.
Rahvuspargi kõrgeim punkt (39 m ü.m.p) asub selle idaosas luiteahelikul. Soomassiivide
vahele jääva mineraalmaa-ala absoluutkõrgus on enamasti 20-25 m vahemikus. Rabade
pealispinna absoluutkõrgus on mõnevõrra suurem: Kuresoo 26-29 m, Ördi raba 26-28 m,
Kikepera 25-28 m ja Valgeraba 25-28 m (Kink, 1996).
Mullastiku poolest kuulub Soomaa kamar-, glei- ja lammimuldade (settealade) Lääne-Eesti
valdkonda, mis jaguneb omakorda kaheks allvaldkonnaks (Lillema, 1958):
· gleistunud kamar-leet-, kamar-glei- ja lammimullad;
· gleistunud kamar-, leet-, kamar-glei- ja lammimullad ning rabamullad.
Jõgede lammidel on levinud lammimullad, mida rühmitatakse järgmiselt:
· lammi-kamarmullad (lühiajaliselt üleujutatavad; teralised, kihilised);
· lammi kamar-gleimullad (lühi- ja pikemaajaliselt üleujutatavad; teralised, kihilised);
· ibejad (mudajad) lammi madalsoomullad (pikemaajaliselt üleujutatavad).
Hüdrograafiliselt kuuluvad Soomaa jõed Liivi lahe vesikonna Pärnu jõgikonda.
16
Soomaa veestiku võib üldjoontes jaotada kahte gruppi:
1. Eesti oludes suhteliselt suured transiitsed jõed;
2. väga tihe mikroveestik, mis koosneb valdavalt metsakuivenduskraavidest.
Vahepealseid, lühikesi ja väikese valglaga jõgesid esineb siin suhteliselt vähe. Peajõgede I ja
II järgu lisajõed esinevad arvukamalt Soomaast itta jäävas piirkonnas (Järvet, 1994). Soomaa
jõed seob ühtseks hüdrograafiliseks süsteemiks Pärnu jõgi, kuhu jõuab vesi peaaegu kõigist
Soomaa jõgedest. Soomaa rahvuspargi territooriumi hüdrograafilise süsteemi suleb Navesti
jõgi, mis on ühtlasi kaitseala põhjapiiriks. Soomaa peajõeks loetakse Halliste jõge koos lisajõe
Raudnaga, mis moodustavad vooluvee peatelje Sakala kõrgustikult Pärnu madalikule (Järvet,
1994).
Soomaa omapäraks on jõgede kõrged ja pikaajalised üleujutused, mis on ainulaadsed kogu
Põhja-Euroopas. Navesti, Halliste, Raudna ja Lemmjõe alamjooksu tuntakse Riisa
üleujutusalana. Halliste jõgi suubub vastuvoolu Navesti jõkke ning põhjustab kevadeti ja
sügiseti suurvee (Allilender, 1997). Üleujutus võib kesta mõnest päevast kuni mõne kuuni.
Suurvee maksimum esineb tavaliselt jäämineku ajal (Allilender, 1997).
Tavalistel aastatel hõlmab üleujutus ca 50 km2 suuruse ala, olles ühest küljest piiratud
Kikepera rabaga, teisest küljest Kuresoo ja kolmandast Öördi rabaga.Viimase saja aasta
jooksul on eriti suured üleujutused on olnud neljal aastal: 1923, 1931, 1951. ja 1956. Kõrgeim
veetase registreeriti 1931. a. 25. aprillil Riisal 21,94 ja Aesoos 21,96 m ü.m.p. See
põhjustas üleujutuse, mis ulatus 175 km2ni (Järvet & Karu 1999).
Antud töös uuriti kaardianalüüsiga majandamise katkemisest tingitud maastikumustri
muutust. Lähema vaatluse alla võeti Soomaa rahvuspargis Halliste luha mõlemal kaldal olev
piirkond (joonis 1), mis administratiivselt jääb peamiselt Pärnu maakonna Tori valla Riisa
külla; lisaks paarihektariline maatükk Viljandi maakonna Kõpu valla Tipu külast.
Taimkatte uuringute eeesmärgiks oli kirjeldada veereziimi ja majandamisviisi mõju
luhataimkatte eluvormilisele struktuurile, liigirikkusele ja maapealsele biomassile.
17
Joonis 1. Kaardianalüüsil kasutatud uurimisala paiknemine ja jaotus.
18
2.2. Materjal ja metoodika
2.2.1. Kaardianalüüsil kasutatud materjal ja metoodika
Puisniitude ajalugu on uuritud erinevate ajalooliste kaartide, aerofotode ning hiljutiste
taimkattekaartide põhjal. Maakasutuse ja taimestiku muutused on omavahel tihedalt seotud
ning kaardiaanalüüsi meetod sobib nende seoste väljaselgitamiseks (Pärtel jt., 1999; Meier,
2001). Eesti maakasutuse ja maastiku muutuste uurimisel on kasutatud lisaks
kaardimaterjalidele (Palang, Mander jt. 1998) maakatastri andmeid (Kasepalu, 1991; Mander
jt. 1994).
Halliste puisniidu keelualal hinnati sealseid taimkatte pindalalisi muutusi. Selleks kasutati
järgmisi kaarte:
1) Eesti Baaskaart
Vastavalt Eesti Baaskaardi lehtede nomenklatuurile kasutatakse digitaalkaardilehti nr 5341 ja
5343, M 1:50 00;
2) "Vene 1-verstane" kaart, M 1:42 000, koost. 1893-1913
Uurimisala paikneb digitaalkaardilehtedel nr 5375 ja 5385;
Togogaafilised kaardid:
3) M 1:50 000 (1905);
4) M 1:25 000 (1947);
5) Botaanilise keeluala "Halliste puisniit" piiridega kaart, M 1:25 000 (1959);
6) Riikliku kaitseala "Halliste puisniit" kaart planeeritavate piiridega, M 1:10 000
(1978);
7) Katastri aluskaart, M 1:10 000, koost. 1985-1986;
19
8) Soomaa rahvuspargi luhtade ja majandamisklasside piiridega kaardid MapInfo
formaadis, koost. 1999;
9) Eesti Põhikaart (digitaalkaart) M 1:10 000, koost. 2002
Kasutatud kaartidest on kõige detailsem 1978. a. koostatud plaan ning katastri aluskaart,
millel on kõlvikute piirid kõige selgemini eristatavamad. Väiksemamõõtkavaliste kaartide
järgi on kõlvikute vahel piire tõmmata veidi keerulisem.
Kaardianalüüsil kasutati MapInfo 6.5 tarkvara. Paberkaardid skaneeriti. Saadud rasterkaart
registreeriti (seoti koordinaatidega) MapInfo keskkonnas vastavalt selgelt eristatavatele
kohtadele rasterpildil ning vektorkaardil Eesti Baaskaardil (põhikaardi projektsioonis).
Antud juhul osutus sobivaks kasutada kaartidel olevaid sarnaseid jõemeandreid.
Eelpool nimetatud kaardimaterjalilt eristati niidu-, võsa ja metsaalasid. Selleks digitaliseeriti
nende alade piirid arvestades kaartidel olevaid leppemärke. Tekitatud polügoonidel võrreldi
erinevate koosluste pindalasid. Nende paremaks võrdlemiseks on võetud põhjapiiriks Soomaa
rahvuspargi välispiir (joonis 2), mis ühtib Halliste puisniidu praeguste piiridega ning lõunas
1959. a. loodud keeluala ulatusega, mille looduslikuks piiriks võib üldjoontes võtta Halliste
jõe, mida nii idast kui ka läänest piiravad metsamassiivid. Samuti pöörati tähelepanu
metsasuse suurenemisele erinevatel maapinna kõrgustel.
Uuritavas piirkonnas hinnati metsakoosluse esinemist alla 20 m absoluutses kõrgusskaalas.
Kasutati 1947. a. topograafilist kaarti (M 1:25 000).
20
Joonis 2. Kaardianalüüsil kasutatud uurimisala Vene 1-verstasel kaardil oleva luhapiiri,
1959.a. loodud keeluala piiri ja Soomaa rahvuspargi välispiiriga.
21
2.2.2. Taimkatte uuringute materjal ja metoodika
Välitööd taimkatte uurimiseks Soomaa rahvuspargi luhtadel viidi läbi 2002. aasta suvel.
Uurimisalad paiknesid (lisa 3) Lemmjõe luhtade piirkonnas Mulgi heinamaal (tähistatud
Mulgi 1, 2), Halliste jõe luhtade piirkonnas Halliste puisniidul ja Halliste keskluhal (Halliste
1, 2, 3), Tõramaa jõe luhtade piirkonnas Tõramaa luhal (Tõramaa 1, 2) ja Raudna jõe luhtade
piirkonnas Kuusekäära luhal (Kuusekäära 1, 2).
Uuriti kuut niiske ja viit märja luhaniidu kasvukohta (tabel 1) (klassifikatsioon Paal, 1997
järgi). Kralli jt. (1980) detailsema klassifikatsiooni järgi kuuluvad uuritud niisked luhaniidud
märgade aasarohumaade ja märjad luhaniidud suurkõrreliste lammirohumaade alla.
Tegelikkuses erinevad nende kahe kasvukohatüübi niiskustingimused vähe. Märjas on
kevadine üleujutus pikaajalisem, seega on ka mulla veega küllastatus kogu
vegetatsiooniperioodil suurem. Meie mõõtmisel oli soojal suvepäeval (26. juunil 2002) mulla
huumushorisondi keskmine veesisaldus niiskes kasvukohatüübis 41,9±5,3% ja märjas
kasvukohatüübis 53,4±9,5% (lisa 4).
Uurimisala luhad jaotati niitude kasutustraditsioone ja majandatavust arvestades järgmiselt
(tabel 1):
1. regulaarselt niidetav ala (A);
2. lühemat aega majandamata ala (B);
3. pikemat aega majandamata (vähemalt 15 aastat niitmata) ala, kus on hiljuti
taasalustatud niitmisega (C);
4. pikemat aega majandamata (vähemalt 15 aastat niitmata) ala, kus on loobutud
niitmisest (D).
Igalt uurimisalalt valiti proovitükk, milleks oli enam-vähem homogeense taimkattega ala.
Antud juhul võib proovitükki võrdsustada taimekooslusega. Proovitüki valikul arvestati
taimkatte mosaiiksust e. laigulisust ja taimkatte kõrgust. Proovitükilt määrati esinevate
taimeliikide arv, mis ühtlasi on tegelik liigifond Pärtel jt. (1996) järgi. Igalt proovitükilt tehti
viis taimkatte analüüsi pandliku prooviruudu suurusega kasutades Zobeli & Liira (1997)
metoodikat. Selle järgi prooviruudu suuruseks ei ole mitte kindel pindala vaid kindla arvu
22
taimevõrsete alla jääv pind. Antud töös kasutasime 500 võsu suurust prooviruutu.
Prooviruutudelt lõigati kõik võrsed liigirikkuse ja maapealse biomassi suuruse määramiseks.
Kokku koguti 55 prooviruudult 26000 võsu.
Laboratoorsed tööd viidi läbi Tallinna Ülikooli Ökoloogia Instituudi laboris, mille teostajaks
oli Laimi Truus. Prooviruutudelt kogutud võsud sorteeriti liigiti ja loendati iga liigi võsude
arv. Arvutati iga liigi võsude % -line osa proovis, mille järgi määrati eluvormide suhe.
Lõpuks proovid kuivatati 48h 70ºC juures ning kaaluti.
Tabel 1. Uurimisalade niiskus- ja majandamisreziimid.
Nr. Uurimisala Kooslusetüüp Niiskusreziim (Truusi & Majandamisreziim
(Paal, 1997 järgi) Tõnissoni, 1998 järgi)
1 Tõramaa 1 Märg lamminiit Regulaarselt üleujutatav, Regulaarselt niidetav (A)
kiiresti tahenev (Märg)
2 Tõramaa 2 Niiske lamminiit Regulaarselt üleujutatav, 15 aastat niitmata (D)
kiiresti tahenev (Niiske)
3 Mulgi 1 Niiske lamminiit Regulaarselt üleujutatav, Regulaarselt niidetav (A)
kiiresti tahenev (Niiske)
4 Mulgi 2 Märg lamminiit Halvasti tahenev kuni Regulaarselt niidetav (A)
liigniiske (Märg)
5 Halliste 1 Niiske lamminiit Regulaarselt üleujutatav, 15 aastat niitmata ala, kus on
kiiresti tahenev (Niiske) hiljuti taasalustatud niitmisega
(C)
6 Halliste 2 Niiske lamminiit Regulaarselt üleujutatav, 15 aastat niitmata (D)
kiiresti tahenev (Niiske)
7 Halliste 3 Märg lamminiit Regulaarselt üleujutatav, 15 aastat niitmata (D)
kiiresti tahenev (Märg)
8 Karusekose 1 Niiske lamminiit Regulaarselt üleujutatav, Lühemat aega majandamata
kiiresti tahenev (Niiske ala (B)
9 Karusekose 2 Märg lamminiit Halvasti tahenev kuni Lühemat aega majandamata
liigniiske (Märg) ala (B)
10 Kuusekäära 1 Niiske lamminiit Regulaarselt üleujutatav, 15 aastat niitmata ala, kus on
kiiresti tahenev (Niiske hiljuti taasalustatud niitmisega
(C)
11 Kuusekäära 2 Märg lamminiit Halvasti tahenev kuni 15 aastat niitmata ala, kus on
liigniiske (Märg) hiljuti taasalustatud niitmisega
(C)
23
Välitöödel kogutud ja kameraalsel analüüsil saadud andmete põhjal saadi järgmised taimkatet
iseloomustavad parameetrid:
Tegelik liigifond (Spool) kõik kooslusest (proovitükilt, mis on silma järgi otsustatult
topograafiliselt ja ökoloogiliselt homogeenne ala) registreeritud taimeliigid;
Liigirikkus 500 võsu kohta (S500) 500 võrse suuruse prooviruudu liikide arv;
Võrsetihedus võsude arv pinnaühiku kohta (võsu/m²);
Pindala500 võrset [m2] muutliku suurusega prooviruudu (500 võrse suurune) pindala;
Ühevõsupindala (PUA) pindala, millel kasvab üks taimevõsu; arvutatud: 500 võsu
pindala/500 (cm²);
Suhteline liigirikkus 500 võsu kohta (Srel500) tegelikust liigifondist 500 võsu suurusel
proovitükil olev osa, arvutatud (Srel.500 = S500/Spool);
Ühevõsubiomass (PUB) biomass ühe võsu kohta: arvutatud: 500 võsu biomass/500;
Maapealne biomass (PUB) kindlalt pinnaühikult kogutud taimede maapealsed osad,
kuivatatud (g/m²).
Eluvormid (Leht, 1999 järgi):
· puhmikulised puhmikuid moodustavad kõrrelised ja lõikheinalised
· võsundilised võsundiliselt levivad kõrrelised ja lõikheinalised
· rohundid liblikõielised ja kaheidulehelised rohundid (suhteliselt laiu lehti
moodustavad taimed)
Statistilisel andmetöötlusel kasutati programmpaketti STATISTICA. Eluvormide jaotumise,
võsude pindala, biomassi ja liigirikkuse seoseid majandamis- ja niiskusreziimiga analüüsiti
kasutades ühefaktorilise dispersioonanalüüsi (one way ANOVA) moodulit. Töötluste vaheliste
erinevuste olulisuse mõõtmiseks kasutati Tukey testi. Tulemused loeti statistiliselt
usaldusväärseks P
Nende alusel on koostatud artikli käsikiri, mis analüüsib lammirohumaade taimkatte
liigirikkuse, bioproduktsiooni ja taimkatte muutuste seoseid niiskus- ja
majandamistingimustega (lisa 4).
24
3. TULEMUSED
3.1. Niidu- ja metsakoosluste pindala muutused 20. sajandil
"Vene 1-verstaselt" kaardilt (joonis 2) võib näha, et niiduala jaguneb oma kujult (kitsenedes
läheb üle järgmiseks osaks) kolmeks terviklikuks luhakompleksiks: põhja-, kesk- ja
lõunaosaks (joonis 1).
1) Põhjaosa
Põhjaosa oli neist kolmest kõige ulatuslikum (ligikaudu 234 ha). Põhjaosa Halliste jõe
vasakul kaldal on põõsastikega heinamaa ca 300-400 m laiuse, kohati ka laiema ribana. Ala
loodeosast arvestatakse välja ca 16 ha suurune ala (joonis 2), mis tähistab "Vene 1-verstasel"
kaardil metsa. Arvutikaardil jääb selle maatüki rahvuspargi territooiumil olev osa
sihtkaitsevööndisse. Halliste ja Lemmjõe vaheline ala on märgitud hõreda puistu ja
põõsastega niidualaks ehk puisniiduks. Puisniiduala on kolmest uurimisala osast siin
pindalaliselt suurim.
"Vene 1-verstasel" kaardi järgi oli niidu- ja metsaalade piiride tõmbamine raskendatud nende
ähmasuse tõttu. 1905. a. kaardil on metsamassiiv niidust selgemini eristatav. 1905. a. kaardil
on kogu ala märgitud hõreda puistuga heinamaaks e. puisniiduks. Niidukooslus on 1905. a.
kaardil samades piirides, kui "Vene 1-verstasel" kaardil.
1947.a. kaardil on kogu luhaala tähistatud samuti puisniiduna. Sellest ajaperioodist ilmnevad
juba olulised erinevused niidu pindala vähenemises (joonis 3) ning leiab aset koosluste
vahetumine e. suktsessioon. Niidukoosluste asendumisest metsaga annavad tunnistust jõe
paremkalda luhal mõnehektarilise pindalaga metsatukad, samuti on märgata metsa pealetungi
jõe vasakkalda kagunurgast.
Paremkaldal on niidukoosluste olukord märksa parem. 1947. aastaks on 20. sajandi algusega
võrreldes paremkaldal metsa osatähtsus suurenenud ca 6% võrra, vasakkalda metsasus on
kasvanud aga üle 20% võrra.
Katastri aluskaardil on puisniidud Halliste jõe mõlemal kaldal asendunud valdavalt alla ühe
hektariliste metsalaikudega. Niidukooslused on vähenenud ca 36% võrra.
25
Eesti Põhikaardil võis eristada looduslikku rohumaad ning metsaala. 20. sajandi lõpuks on aga
niidukooslused vähenenud sajandi algusest ca 65% võrra.
250
200
150
Pindala ha
Mets
Niit
100
50
0
1900 1920 1940 1960 1980 2000 2020
Aasta
Joonis 3. Metsa- ja niidukoosluste pindala muutused (1905-2002) uurimisala põhjaosas.
2) Keskosa
Uurimisala keskosas on luhapindala 1905. a. kaardil ca 162 ha. Luha laius kitsamates
kohtades on 200-300 m, laiemates kohtades ulatub 600-700 meetrini. "Vene 1-verstasel"
kaardil ning 1905. a. kaardil on see ala tähistatud hõrendikuga heinamaana.
1947. a. kaardilt ilmneb niiduala vähenemine (joonis 4) kogu keskosas ca 20% võrra.
Katastri aluskaardi järgi on niidupindala sajandi algusest 1985-1986ndateks vähenenud 51 %
võrra. Kaardilt võib eristada paremkaldal kaht erinevat niidukooslust: hõrendikuga heinamaad
e. puisniitu (ca 30 ha) ning ainult heinamaana (ca 15 ha) tähistatud kõlvikuid. Metsamaa
pindala on ca 40 ha. Vasakkaldal on puisniitu säilinud ca 15 ha, heinamaad on ca 30 ha ning
ca 35 ha on tähistatud metsamaana.
26
20. sajandi algusest on niidukooslusest säilinud sajandi lõpuks vaid 7%; 93% on kattunud
metsaga.
180
160
140
120
Pindala ha
100
Mets
80 Niit
60
40
20
0
1900 1920 1940 1960 1980 2000 2020
Aasta
Joonis 4. Metsa- ja niidukoosluste pindala muutused uurimisala keskosas (1905-2002).
3) Lõunaosa
Uurimisala lõunaosa Vodi luhal ja Tõramaa puisniidu alal tähistavad "Vene 1-verstasel"
kaardil ja 1905. a. kaardil põõsad ja hõre mets puisniitu. 20. sajandi algul oli luhapindala ca
62 ha. Paremkalda luha laius oli 300-400 m, vasakkalda luha laiuseks võib lugeda 500-800 m.
Nii nagu põhja- ja keskosas, on ka lõunaosas toimunud niidukoosluse pindalaline vähenemine
(joonis 5) ja metsasuse kasv. Paremkalda metsa pealekasv servaaladelt on olnud 20. sajandi
algusest kuni sajandi keskpaigani ca 30%. Vasakkaldal on ala sees metsatukk, mis moodustab
ca 1% ala pindalast.
Kogu uurimisala lõunaosast on 20. sajandi keskpaigaks metsaga kaetud ca 10%.
27
Katastri aluskaardil olevast niidualast on paremkalda luhast 50% kattunud metsaga,
vasakkaldal on metsasus alla 40%. Kogu lõunaosas on luhakooslusest säilinud sajandi
algusest 1985ndateks ca 60%.
Sajandi jooksul on lõunaosas niidukooslus vähenenud sajandi lõpuks 75% võrra.
70
60
50
Pindala ha
40
Mets
Niit
30
20
10
0
1900 1920 1940 1960 1980 2000 2020
Aasta
Joonis 5. Metsa- ja niidukoosluste pindala muutused (1905-2002) uurimisala lõunaosas.
Tabel 2. Uurimisala põhja-, kesk- ja lõunaosa niidu- ja metsakoosluste ligikaudsed pindalad
(ha) ajavahemikus 1905-2002.
Põhjaosa Keskosa Lõunaosa Kogu uurimisala
Aasta
Niit Mets Niit Mets Niit Mets Niit Mets
1905 ~234 0 ~162 0 ~61 0 ~457 0
1947 ~199 ~35 ~130 ~32 ~55 ~6 ~384 ~73
1985 ~149 ~85 ~80 ~82 ~35 ~26 ~264 ~193
2002 ~81 ~153 ~12 ~150 ~15 ~46 ~108 ~349
28
Kokkuvõtvalt võib öelda, et üldised trendid kogu uurimisalal ja selle erinevatel osadel (tabel
2; joonis 6) ajavahemikus 1905-2002 on:
1. luhapindalade järk-järguline vähenemine;
2. pindalade vähenemise kiirenemine pärast 1950ndaid.
90
80
70
60
Põhjaosa
50
Keskosa
%
40 Lõunaosa
Kogu uurimisala
30
20
10
0
1940 1950 1960 1970 1980 1990 2000 2010
Aasta
Joonis 6. Uurimisala põhja-, kesk- ja lõunaosa niidukoosluse pindalaline vähenemine %-des
ajavahemikus 1905-2002.
Niidukoosluste pindalaline vähenemine on toimunud järgmiselt (tabel 3):
1. Ajavahemikus 1905-1947 on pindala vähenenud uurimisala põhja-, kesk- ja lõunaosas
15-20%, kogu uurimisalal 16%;
2. 1985daks aastaks on uuritavas piirkonnas luhapinadala kahanenud 25-38%, kogu
uurimisalal 31%.
3. 2002 aasta Põhikaardi järgi on looduslike rohumaade pindala vähenenud 1985ndatest
põhjaosas 45%, keskosas 85% ja lõunaosas 57%, kogu uurimisalal 59%.
29
Tabel 3. Luhakoosluste pindala (%) vähenemine uurimisala põhja-, kesk- ja lõunaosas
erinevatel ajavahemikel.
Ajavahemik Põhjaosa Keskosa Lõunaosa Kogu uurimisala
1905-1947 ~15% ~20% ~10% ~16%
1947-1986 ~25% ~38% ~36% ~31%
1986-2002 ~45% ~85% ~57% ~59%
Luhapindala vähenemine on toimunud kõige kiiremini ja muutused on olnud drastilisemad
uurimisala keskosas (joonis 6). Põhjaosas on see protsess toimunud teiste osadega võrreldes
veidi aeglasemalt. Niiduala on uurimisala osadel hakanud vähenema järsemalt peale 1947.
aastat (tabel 3). 20. sajandi algusest on sajandi lõpuks kogu uurimisala niidukooslusest
säilinud ca 24% ja metsasus on suurenenud ca 76% võrra.
3.2. Looduskaitseliste meetmete rakendamise mõju uurimisala luhakoosluste pindalade
levikule
1959.a. kaitse alla võetud territooriumi (lisa 2) suurus on erinevate andmete järgi veidi erinev.
Kirjanduse (Kumari, 1960) järgi on see ca 265 ha, 1959 a. kaardi järgi hinnates ca 274 ha.
1978. a. kaardistati kõnealune piirkond Eesti metsakorralduskeskuse poolt. Koostati kaitseala
plaan, kirjeldus ning tehti majandamisettepanekud. Kaardi järgi on projekteeritud piiridega
keeluala pindala 245 ha.
1959. ja 1978. a kaartidel on kaitseala piiridesse jäänud ka osa metsaalast. Kaitse alla ei ole
võetud kogu säilinud niiduala. 1978. a kaardil on hõreda niidu ja põõsastega puistut või
heinamaad 236 ha. Nii 1959. a. kui ka 1978. aasta kaartidel esineb kaitstava territooriumi
piiride tõmbamisel palju ebatäpsusi. Keeluala piiride määramisel ei ole arvestatud seda, et piir
jookseks mööda luhakõlvikute piire. Kohati jääb sinna metsakooslust, mis pole seotud
kaitseala loomise eesmärkidega. 1959. a. botaanilise keeluala piir (lisad 1, 2) tõmmati
põhimõttel ,,200 m mõlemale poole jõe kaldast", kuid seda reeglit pole väga rangelt järgitud.
Kaardimaterjali põhjal võib öelda, et uurimisala põhja- ja keskosas ulatub 1959. a. keeluala
luha laius ligi 300-400 meetrini, lõunaosas 300-600 meetrini. 1959. a. loodud botaanilise
keelualast (265 ha) on 2000. aastaks sisuliselt säilinud 27 ha. 1959. a. kaitse alla võetud
30
luhakooslusest on hästi säilinud ca 10%. Kaitseala loomisega ei suudetud pidurdada
niidukoosluse pindalalist kahanemist.
Kaitsekorralduskavas (Allilender, 2000) on Halliste jõe luhad jagatud majandamisklassidesse.
1. majandamisklassi kuuluvatel rohumaadel toimub niitmine igaaastaselt. 2. klassi rohumaid
niidetaks vähemalt üle aasta ja 3. klassi niite tuleb niita vähemalt üle kolme aasta. 4. klassi
luhtadel niitmist ei toimu, kuna need on tugevalt võsastunud ning võimalusel tegeletakse
võsakoristusega. Igal aastal niidetavat luhta (1A kl.) on uurimisala põhjaosas kokku 12 ha ja
lõunaosas Tõramaa puisniidul 15 ha. Uurimisala niidud kuuluvad 1., 2., 3. ja 4.
majandamisklassi niitude hulka. Keskosas on valdavalt 3. ja 4. majandamisklassi luhaniidud
(Suurkask, 1999.)
Rahvuspargi andmetel on kogu rahvuspargi territooriumil lisaks hooldustöödele hakatud
tegelema ka luhaniitude taastamisega. Aastatel 2001-2005 on taastatud luhaniite kokku ca 255
ha.
Viimastel aastatel on rahvuspargis taastatud ja majandatud luhaniitude pindala (joon. 7) veidi
kasvanud. Hooldustöid teostatakse vastavalt sellele, kuidas riigipoolselt töid finantseeritakse.
Luhtade majandamiseks on korraldatud ka talguid alates 1994. Aastast. Põhiliselt on need
toimunud kohalike rahvuspargi töötajate (Tõnu Kütt, Kaja Allilender, Meelis Suurkask,
Gunnar Sein jt.) kaasabil. Sagedasemad talgulised on olnud Soomaa Sõprade Selts ja
Pärandkooslute Kaitse Ühing.
31
450
400
350
300
Pindala ha
250
200
150
100
50
0
2000 2001 2002 2003 2004 2005
Aasta
Joonis 7. Soomaa rahvuspargi luhaniitude hooldustööd aastatel 2000-2005.
Euroopa Liidu LIFE-Nature projekti (EAGELLIFE) raames on hakatud luhti taastama
laiemal maa-alal kui kunagi varem. Projekti kestuseks on 01.04.2004 - 31.05.2009. Selle
käigus taastatakse rahvuspargis erinevas kvaliteedis Natura 2000 biotoope (luhti) 1994
hektaril. Projekti käigus on seni taastatud põhjaosas (Halliste puisniidul) 33 ha, keskosas
(Halliste keskluhal) 98 ha, lõunaosas (Tõramaa puisniidul) 22 ha. Taastamist plaanitakse
jätkata kogu rahvuspargi territooriumil olevatel luhtadel.
32
3.3. Uurimisala metsasus erinevatel maapinna kõrgustel
Võrreldes omavahel 1947. a. topograafilist ja 1985. a. katastrikaarti, selgusid järgmised
muutused metsade pindalas:
1) põhjaosa
1947. a. kaardil on põhjaosa metsaalast 15% alla 20 m absoluutses kõrgusskaalas. Katastri
aluskaardil on selleks näitajaks 3%.
2) keskosa
1947. a. kaardil on metsasus madalamal kui 20 m absoluutkõrgusel ca 16% kogu seal
paiknevast metsast. Katastri aluskaardil on 20% kogu keskosa metsakooslusest madalamal
maapinna kõrgusel.
3) lõunaosa
1947. a. kaardil on ca 7% kogu metsakooslusest madalamal kogu lõunaosa metsapindalast.
Katastri aluskaardil ei ole metsa pindala madalamal maapinna kõrgustel muutunud.
Uurimisala keskosas on madalamatel aladel oleva metsa osatähtsus kogu metsaalast veidi
suurem (tabel 4). Olemasolevat kaardimaterjali kasutades võib öelda, et valdavalt on metsasus
suurenenud kiiremini kõrgematel aladel ning mets tungib peale metsa servadelt. Madalamal
maapinna kõrgusel toimub metsastumine aeglasemalt.
Tabel 4. Metsa osatähtsus madalamatel maapinna kõrgustel uurimisala põhja-, kesk- ja
lõunaosas.
Aasta
Uurimisala osa
1947 1985
Põhjaosa 15% 3%
Keskosa 16% 20%
Lõunaosa 7% 0%
33
3.4. Niiskustingimuste ja majandamisreziimi mõju luha taimkattele
Uuritavate luhtade iseloomulikeks liikideks märja luhaniidu kooslusetüübis on niiske
luhaniidu kooslusetüübis Deschampsia caespitosa (luht-kastevars), Festuca rubra (punane
aruhein), Filipendula ulmaria (harilik angervaks) ja Deschampsia cespitosa (luht-kastevars),
Carex cespitosa (mätastarn), Calamagrostis stricta (püstkastik).
Taimkatte liigirikkuse ja biomassi seoseid niiskustingimuste ja niitmisreziimiga on
detailsemalt esitatud lisas 4. Andmed proovipindala, maapealse biomassi suuruse, liigirikkuse
ja liikide eluvormilise jaotuse kohta niisketes ja märgades koosluse tüüpides on toodud tabelis
5 ja nende näitude statistiline olulisus tabelis 6. Võrsetihedus (m2 kohta), ühevõsupindala ja
ka paindliku suurusega prooviruut ei erinenud statistiliselt oluliselt erinevate
niiskusreziimidega prooviruutudel. Ka maapealse biomassi suurus ei olnud statistiliselt
oluliselt sõltuv niiskustingimustest. Liigifond oli suurem niiskes kasvukohas, kuid 500 võsu
suuruse prooviruudu liigirikkus ei sõltunud niiskustingimustest.
Majandamisreziimidest eristus kõige selgemini reziim D, mis erines ülejäänutest
võrsetiheduse ja võrsepindala suuruste poolest (Anova test). Nendel näitajatel ei olnud
regulaarselt niidetud (reziim A), kui ka osaliselt niitmata alade (reziimid B ja C) vahelised
erinevused olulised. Spool ja S500 näitajad eristasid reziimid A ja B reziimidest C ja D. Kuigi
reziimil C ei olnud võrsed oluliselt suuremad kui reziimidel A ja B, olid nii liigifond kui
prooviruudu liigirikkus statistiliselt oluliselt väiksemad.
Eluvormidest olid rohundid enam levinud (% võsude arvust) niiskes kasvukohas, kui ei
niidetud regulaarselt (reziimid B, C, D). Märjas kasvukohas regulaarselt niidetavatel aladel
(reziim A) ning pikaajaliselt niitmata (reziim D) domineerisid puhmikulised. Võsundilised
domineerisid ebaregulaarse majandamisega aladel (reziimid B ja C) (tabel 5).
Maapealsel biomassil (500 võsu kohta) ei esinenud statistiliselt olulist seost niiskusereziimi
suhtes (F=0.41, p=0.06), seevastu ilmneb statistiliselt oluline seos majandamisreziimi suhtes
(F=7.91, p=0.00). ANOVA näitas, et biomass 1 m² kohta ei olenenud ei niiskustingimustest
ega ka majandamisreziimist. Kui aga arvutati muutuva suurusega prooviruudu suuruse järgi,
oli majandamisreziimist tingitud erinevus statistiliselt oluline (tabel 6).
34
Maapealne biomass oli väikseim regulaarselt niidetavas (reziimiga A) märjas
kasvukohatüübis (263±108 g/m²) ja kõrgeim majandamata niiske kasvukohatüübis koosluses
(763±627.5 g/m²) (tabel 5).
Liigifond (Spool) oli statistiliselt oluliselt kõrgem (p
(33±6.2) kasvukohtades (tabel 3). See oli aga erinevate majandamisreziimide suhtes
statistiliselt erinev (F=7.05, p
6).
PUA suurenes statistiliselt oluliselt ja võrsete tihedus vähenes, kui majandamine katkes 15
aastaks. Väikseim PUA oli niiskes kasvukohatüübis majandamisereziimi B juures. B- ja C
reziimid erinesid A-reziimist vähe (tabel 5). Niiskusreziimi ja PUA vahel puudus statistiliselt
oluline seos (tabel 6).
35
Tabel 5. Arvutatud liikide koosesinemise ja biomassi jaotuse parameetrid; keskmised koos standardveaga (sulgudes).
Niiskus- ja Proovi
majandami Maapealne ruudu Eluvormide jaotus (% võsude arvust)
Võrsetihedus 2
Pindala500võrset
s- PUA [cm ] biomass Spool liigi-
[võsu/m2] [m2] 2
reziim [kuivkaal g/m ] rikkus Puhmiku-
Võsundilised Rohundid
lised
(vt tabel 1) (500
Märg; A 2321 (845) 5.5 (2.76) 0.276 (0.134) 263 (108.0) 37 23 (3.6) 49 (19.3) 33 (25.7) 18 (16.1)
Märg; B 1012 (247) 10.4 (2.80) 0.520 (0.140) 516 (165.9) 31 12 (8.6) 27 (32.6) 67 (16.7) 6 (12.1)
Märg; C 2142 (1983) 7.5 (4.60) 0.378 (0.231) 452 (398.7) 40 18 (2.0) 19 (14.8) 66 (12.7) 15 (6.2)
Märg; D 708 (665) 25.2 (17.7) 1.258 (0.883) 447 (86.4) 16 5 (4.1) 82 (8.7) 1 (1.6) 17 (9.4)
24
Niiske; A 3468 (1751) 3.4 (1.56) 0.174 (0.080) 572 (692.3) 57 59 (22.9) 25 (15.1) 16 (9.9)
(10.2)
Niiske; B 5024 (2179) 2.6 (1.85) 0.321 (0.067) 333 (167.5) 69 20 (2.1) 24 (28.7) 34 (18.5) 43 (11.2)
Niiske; C 3194 (1174) 5.3 (1.71) 0.397 (0.605) 380 (178.2) 36 17 (2.4) 33 (3.7) 44 (14.1) 24 (12.6)
Niiske; D 745 (175) 17.0 (6.35) 0.815 (0.309) 763 (627.5) 41 13 (6.1) 25 (23.2) 31 (32.1) 44 (53.7)
Tabel 6. Peamiste taimeliike iseloomustavate näitajate kleskmised niiskustingimuste järgi (A) ja niitmisreziimi järgi (B) koos ANOVA
tulemustega. Numbrid on keskmised koos standardveaga (sulgudes). Statistiliselt olulised seosed (p
A
Parameeter Niiskustingimused ANOVA tulemused
Märg Niiske F P
Võrsetihedus [võsu/m2] 1701 (1263)a 2209 (1977)a 1,23 0,273
PUA [cm2] 10,8 (10,9)a 9,0 (8,2)a 0,36 0,55
Pindala500võrset [m2] 0,523 (0,540)a 0,462 (0,433)a 0,36 0,55
Ühevõsubiomass [g] 0,5 (0,59)a 0,8 (2,03)a 0,41 0,53
Liigifond 33a 46a 12,75 0,0009
Prooviruudu liigirikkus 16 (8,1)a 19 (8,3)a 0,906 0,346
Puhmikulised [%] 42 (14,1)a 37 15,2)b 8,072 0,006
Võsundilised [%] 44 (20,3)a 29 (12,5)a 0,101 0,752
Rohundid [%] 14 (6,5)a 33 (17,0)b 19,641 0,000
B
Parameeter Majandamisreziim ANOVA tulemused
A B C D F P
Võrsetihedus [võsu/m2] 2703 (1337) a
1518 (1042)a 2360 (2127)ae 586 (479)bf 4,47 0,007
PUA [cm2] 4,8 (3,07)ace 8,4 (3,44)ace 7,1 (3,84)ace 24,9 (13,85)bdf 21,317 0,000
Pindala500võrset [m2] 0,242 (0,153) ace
0,419 (0,171)ace 0,536 (0,542)ace 1,245 (0,691)bdf 2,331 0,000
PUB [g] 0,1 (0,12)ace 0,4 (0,23)ace 0,2 (0,14)ace 2,5 (3,12)bdf 7,919 0,0002
Spool 46ace 50ace 37ade 29bdf 7,048 0,0004
acf
S500 23 (6,2) 21 (10,6)ace 16 (3,9)bce 9 86,7)bdf 10,374 0,000
Puhmikulised [%] 52 (10,4)ade 44 (27,9)bcf 36 (14,7)ade 55 (28,9)ade 5,255 0,003
ade
Võsundilised [%] 30,1 (12,0) 50,6 (17,4)bcf 40,7 (10,0)ade 6,1 (5,5)bdf 14,246 0,000
Rohundid [%] 17 (2,3)ace 24 (19.5)ace 23 (4,7)ace 39 (23,9)bcf 4,534 0,007
4. ARUTELU
4.1. Kaardianalüüsi tulemused
Kaardianaüüsil ilmnes niidu pindala järk-järguline kahanemine ja metsasuse suurenemine.
Kaardimaterjali kesisus, mõõtkavade erinevus ja ebaühtlane vanus raskendasid andmete
võrdlemist.
Antud uurimistööks vajalikku erinevatest ajaperioodidest pärinevat kaardimaterjali õnnestus
arhiividest leida ebapiisavalt. Taimekoosluste pindala muutuste jälgimiseks oleks olnud hea,
kui kaardid oleksid olnud mingite kindlate ajavahemike tagant, nt iga kümne aasta järel.
Sellisel juhul oleks olnud lihtsam leida põhjuseid ja seoseid koosluste pindala muutuste kohta.
Samuti oleks olnud võimalik hinnata, kui kiiresti niidud majandamise lakkamisel
metsastuvad.
Kaardimaterjali mõõtkavade erinevuste tõttu ei ole võimalik anda väga täpseid arve. On
toodud vaid ligikaudsed pindalad ja üldised trendid.
Hetkel võime näha vaid olukorda, mis esines mõisaperioodi lõpul, kolhooside kõrgajal ning
looduskaitseliste meetmete rakendamisel. Samuti ilmnes metsasuse suurenemise olenevus
maapinna kõrgusest madalamad reljeefiosad metsastusid aeglasemalt.
Antud materjalilt oli võimalik eristada metsa ning niidu- ja puisniidualasid, kuid mitte võsa
suurenemist. Katastri aluskaardilt oli võimalik eristada ka erinevaid niidukooslusi, mis
varasematel kaartidel ei eristunud.
Ala loodenurgas on ca 16 ha suurune maatükk märgitud 20. sajandi alguse kaartidel
metsamaana, hiljem on aga kasutusele võetud heinamaana. (1947. a. kaardil on tähistatud see
ala hõreda metsaga heinamaana.) Võrreldes uurimisala muu osaga on selle ala luhakooslus
suhteliselt noor ning ei oma sellist ajaloolist väärtust kui ülejäänud luhaalad.
38
4.1.1. Halliste luha põllumajanduslikust maakasutusest
Pool-looduslike koosluste leviku kõrgaeg oli Eestis 19. sajandi lõpul, kui looduslikud
rohumaad võisid hõlmata ligikaudu 1/3 maismaa pindalast. Ligilähedane olukord kestis
1940ndateni (Kukk, Kull 1997). Käesolevas töös kasutatud vanimad allikad "Vene 1-
verstane" kaart ning 1905. a. topograafiline kaart näitavad ilmselt kasutuses olnud
niidupindala maksimaalset seisu. Kogu niidukoosluse pindala suuruseks hinnati ca 457 ha
(tabel 2). Halliste puisniitu on heinamaana kasutatud arvatavasti aastatuhandeid.
Ümbruskonna asulate heinamaad asusid enamasti Halliste jõe kaldail, seda kuni 1950.-
1960ndateni. Sellest annab tunnistust ka matkakirjeldus 1960ndatest: "Kahel pool jõge,
niikaugele kui silm ulatub, aina küünid, lausa küünide külad (Tamm, 1967)."
Eestis vähenes pool-looduslike rohumaade pindala 1950ndate aastate 1,5 miljonilt hektarilt
1990. aastaks 0,3 miljonile hetarile (Mander, 1994). 1950ndatel oli Halliste luht sealsete
kolhooside tähtis loomasöödabaas. Näiteks sai Viljandi rajooni "Valguse" kolhoos kogu
sööda sealsetelt luhaniitudelt. Halliste luhaniitu hinnati keskmise majandusliku väärtusega
luhaniitude hulka. Kuid juba sel ajal oli oht niidualade võsastumiseks peamiselt jõega
piirnevatel aladel (Mikk, 1957). Peale 1950daid aastaid on uuritavas piirkonnas
niidukoosluste pindalaline vähenemine kasvanud.
Vasakkalda luht oli 1950ndate lõpul tugevasti võsastunud ning jäi suures osas majanduslikult
täiesti kasutamata, ent paremkaldal levis väga ilus puisniit. Märgitakse, et Halliste puisniidul
jäi hein sageli tegemata suviste üleujutuste tõttu (Pullisaar, 1959).
1970.-1980. aastatel vähenes niidetava maa pindala tunduvalt ning niitmine muutus
ebaregulaarseks. 1981-1982. a. kohta märgitakse, et hõre puurinne võimaldab heina niita
mehhaniseeritult. Puude katvuseks hinnatakse ligikaudu 10%. Sõltuvalt kohalike majandite
söödavajadusest jäi igal aastal osa niitmata, kuid ala seisund oli sellegipoolest rahuldav.
Viimasena niitis Kõpu kolhoos jõe paremkaldal paiknevaid maid heinaniidukitega. Seetõttu
olid need alad paremas olukorras, kuigi varem Halliste puisniitu iseloomustanud üksikud
vanad tammed olid ümbritsetud põõsastest ning puisniidu ilme kannatas tublisti. Botaaniliselt
huvitavamad, kõrgemad kaldaosad, olid kas võsastunud või võsastumas niitmise lõppemise
tõttu (Sarv, 1984). 1985-1986ndateks, kolhoosiaja kõrgajaks, oli niidukoosluse pindalaks ca
264 ha, sajandi algusest oli niiduala vähenenud ca 42% võrra.
39
4.1.2. Looduskaitseliste meetmete rakendamise mõju uurimisala luhakoosluste pindalade
levikule.
Juba 1976. aastal märgiti, et viimase 15-20 aasta jooksul on puisniidud peaaegu kadunud ka
kaitsealadelt. Tehti ettepanek valida välja esinduspuisniidud ja asuda neid vajalikul viisil
hooldama s. t. igal aastal niitma (Sarv, 1984).
1996. a. toimus Halliste luha inventeerimine, et välja selgitada luha kõige väärtuslikumad
osad ning koostada kava edasiseks majandamiseks. Inventeerimise tulemustena selgus, et
Halliste puisniidu säilinud osa suurus on 23 ha, mis oli parimas seisundis, Tõramaa puisniidul
(21 ha) (Suurkask, 1996).
Halliste puisniidu korrashoiuks on Soomaa rahvuspark korraldanud alates 1994. a. mitmeid
talguid. Riikliku Looduskaitsekeskuse seire ja teadustöö spetsialisti Gunnar Seini suulistel
andmetel pole talgute raames tehtavate tööde maht olnud kuigi suur. Selliselt suudetakse
taastada või hooldada vaid mõned hektarid ning antakse suhteliselt väike panus luhtade
taastamisele või säilitamisele. Aasta-aastalt on tõusnud riigipoolne rahaline toetus luhtade
hooldamiseks, vahendeid selleks saadakse veel Euroopa Liidu keskkonnaprogrammi LIFE
kaudu.
Uurimisala erinevaid osi võrreldes, on keskosas toimunud metsastumine kiiremini ilmselt
seetõttu, et seda osa ei ole peetud looduskaitselistest seisukohast nii väärtuslikuks, kui Halliste
puisniitu põhjaosas või Tõramaa puisniitu lõunaosas, mis kuuluvad 1A majandamisklassi
niitude hulka ja mille majandamine toimub regulaarselt. Uurimisala keskosa luhaniidud
kuuluvad valdavalt 4. majandamisklassi luhaniitude hulka (81 ha) (Suurkask, 1999).
40
4.2. Luhaniitude taimkatte uuringute tulemused
Uuritud taimekoosluste struktuuri (liigifond, prooviruudu liigirikkus) ja taimevõsude suuruse
(pindala, biomass) parameetrid seostati kasvukoha niiskustingimustega ja rakendatud
niitmisreziimidega.
Et valisime kaks suhteliselt lähedaste niiskustingimustega kasvukohatüüpi, ei olnud
maapealne biomass ega liigirikkus neis oluliselt erinevad. See ei ole kooskõlas eelnevate
andmetega (Prach, 1992; Touzard jt. 2002). Tulemusi tasandas kindlasti ka võrsete suurust
arvestava (500 võrset) prooviruudu kasutamine. See muutis erineva suurusega taimevõsudega
prooviruudud omavahel paremini võrreldavateks. Fikseerimata suurusega prooviruudud olid
725 kuni 9 900 cm2.
Majandamisreziim mõjutas oluliselt biomassi, liigilise mitmekesisuse ja võrsete tiheduse
näitajaid. Falinska (1991; 1995) leidis samuti, et liigirikkus ja võrsetihedus vähenesid,
maapelne biomass suurenes ja taimevõsude kõrgus ja ühevõsupindala suurenesid niitmise
katkemisele järgneva suktsessiooni käigus.
Eluvormiliselt koosseisult olid sarnased regulaarselt majandatav (reziim A) ja pikka aega
majandamata (reziim D) luhaniidud. Tegelikut kasvasid neil erinevad liigid, mis olid ka
erineva suurusega. Reziimil A domineerisid madalakasvulised (rohurinde alumises osas)
väikesed puhmikuid moodustavad kõrrelised (nt harilik lubikas, lamba-aruhein ), reziimil D
aga suuri kõrgeid puhmikuid (mättaid) moodustad tarnad (mätastarn, luhttarn). Ka sama liigi
puhul (nt. luht-kastevars) olid võrsed ja puhmikud regulaarselt niidetud kohtades palju
väiksemad kui niitmata kohtades. Meie uurimuse tulemustest (lisa 4) nähtub, et juba
lühiajalise majandamise katkemise (mõni aasta) järel hakkab koosluses toimuma muutus nii
liigilises koosseisus kui ka võsude suuruses. Vastupidisel juhul, kui pikka aega majandamata
ja muutunud struktuuriga niidul taastatakse niitmine, toimub muutus rohustu struktuuris
samuti kiiresti. Uuritud taimeliikide parameetrid olid reziimide D ja B vahel palju sarnasemad
kui D ja C vahel.
41
KOKKUVÕTE
Antud töö eesmärgid olid järgnevad: 1) anda luha kui Soomaa ühe võtmeökosüsteemi
iseloomustus; 2) anda ülevaade luhakoosluste hüdroloogilistest ja troofsustingimustest ning
nende seosest majandustegevusega; 3) kaardimaterjali analüüsi rakendades uurida, kuidas on
looduskaitse meetmed ning muutused põllumajanduslikus maakasutuses mõjutanud uuritava
piirkonna luhaniitude pindala; 4) selgitada välja, millised on majandamise katkemise järel
taimkattes toimunud muutused; 5) leida seoseid luhataimkatte struktuuri, maapealse biomassi,
liigirikkuse ja eluvormide jaotumise vahel erinevates niiskustingimustes ja
majandamisreziimidel.
Kaardianalüüsi rakendati Soomaa rahvuspargi Halliste luha ja selle eri osade niitude leviku
dünaamika uurimisel ajavahemikus 1905-2002. Üldine trend kogu uurimisalal kui ka selle
osadel (Halliste luha põhja-, kesk ja lõunaosa) on olnud luhaniitude pindala järk-järguline
vähenemine metsastumise tõttu. Metsastumise protsessi kiirenemine on kaartidelt nähtav
alates 1947. aastast. Valdavalt on metsasus suurenenud kõrgematel reljeefiosadel ning mets
tungib peale metsa servadelt.
1959. aastal loodi botaaniline keeluala "Halliste puisniit" ja rakendati sellel alal esimest korda
looduskaitselisi meetmeid. Kaitseala loomisega ei suudetud pidurdada selles piirkonnas
luhaniitude kinnikasvamist metsaga. 20. sajandi jooksul vähenes kõneallolevas piirkonnas
niidukooslus 76% võrra. Viimastel aastatel on Soomaal hakatud tegelama luhtade
taastamisega. Euroopa Liidu Life-Nature projekti raames planeeritakse taastada rahvuspargi
territooriumil 2009daks aastaks 1994 ha luhaniite.
Luhaniitude taimkatet uuriti Soomaa rahvuspargi kahel suhteliselt lähedase niiskusreziimiga,
kuid erineva majandusreziimiga luhaniidu kasvukohatüübis kokku 11 luhaniidukoosluses.
Maapealne biomass (500 võsu kohta) ei sõltunud statistiliselt olulist niiskusreziimist kuid
sõltus majandamisreziimist. Taimede suurus (PUA; PUB) kasvas statistiliselt oluliselt ja
võrsetihedus vähenes, kui niitmine lakkas 15 aastaks. Niiskusreziimi ja PUA vahel puudus
statistiliselt oluline seos. Liigifond oli statistiliselt oluliselt kõrgem niisketes kui märjemates
kasvukohtades. See oli aga erinevate majandamisreziimide suhtes statistiliselt erinev
liigifond kahanes reziimist A reziimi D suunas.
42
Ebaregulaarselt niidetud aladel (B-, C-reziimiga) ja pikka aega niitmata (D-reziimiga) aladel
domineerisid rohundid. Võsundilised olid rikkalikult esindatud ebaregulaarse reziimiga
aladel. Regulaarselt niidetavatel aladel kui ka pikaajaliselt niitamata aladel domineerisid
puhmikulised kuid need erinesid nii liikide kui suuruse poolest. Regulaarse niitmise korral
olid nendeks alumises rohurindes kasvavad kõrrelised, niitmata aladel aga suuri puhmikuid
moodustavad liigid nagu luht-kastevars, mätastarn.
43
SUMMARY
Changes of the plant cover in the Halliste meadow
The aims of the present thesis were the following: to characterize flood-plain meadow that is
one of the Soomaa key ecosystems; give a survey of the hydrological and trophic conditions
of flood-plain meadow communities and their connections with economic activities; applying
map material analysis to study how the nature protection measures and changes in agricultural
land use have influenced the area of flood-plain grasslands in the studied area; to find out
about the vegetation changes of flood-plain grasslands due to the cessation of management; to
find connections between flood-plain vegetation structure, standing biomass, species richness
and life-form distribution in different moisture and management regimes.
Map analysis was applied in studying Halliste flood-plain meadow and the distribution
dynamics of its different meadows in Soomaa National Park during the years 1905-2002. The
general trend on the whole studied area and on its parts (the northern, middle and southern
parts of Halliste flood-plain meadow) has been the gradual decrease of flood-plain meadow
due to forestation. Predominantly, forestation has increased on higher parts of relief and
advanced at the forest edges.
In 1959 the botanic reserve `Halliste wooded meadow' was founded, and in this region the
nature protection methods were applied for the first time. The foundation of the reserve could
not restrain the overgrowth of flood-plain grasslands with forests in the area. In the 20th
century the grassland community in the particular region decreased by 76%. Within the frame
of the European Union programme LIFE-Nature, 1994 hectares of flood-plain meadows are
planned to be restored in Soomaa National Park by the year 2009.
Flood-plain grassland vegetation was studied in Soomaa National Park on two relatively
similar moisture regime but different management regime flood-plain grassland community
types, in total in 11 flood-plain grassland communities. Standing biomass (per 500 ramets)
did statistically not greatly depend on moisture regime, but on management regime. The size
of plants (PUA; PUB) grew substantially according to statistics, and ramet density decreased
when mowing stopped for 15 years. There was no essential statistic connection between
moisture regime and PUA. Statistically, species pool was substantially higher in moist and
44
wetter community types. But statistically it was different in different management regimes
species pool decreased from regime A towards regime D.
In the unsystematically mowed areas (B and C regime), and in areas that had not been mowed
for a long time (D regime), dominated herbs. Mat-forming graminoids were abundantly
represented in the unsystematic regime areas. Both on regularly mowed areas and long-time
unmown areas dominated tussock-forming graminoids, but they differed for species and size.
In case of regular mowing they were graminaceous growing in lower herb layer, in unmown
areas they were species that form large dwarf shrubs like tussock grass and tussock sedge.
45
TÄNUAVALDUSED
Töö autor tänab prof. Tõnu Oja magistritöö ja kaarditöötluse juhendamise eest. Eriline tänu
kuulub veel MSc. Laimi Truusile, kes oli suureks abiks ning kellel jagus kannatust nii töö
juhendamisel kui ka välitööde praktiliste ja teoreetiliste küsimuste lahendamisel. Samuti tänan
Elle Roosalustet heade nõuannete ja soovituste eest töö osas. Tänan ka Riikliku
Looduskaitsekeskuse töötajaid Gunnar Seini ning Meelis Suurkkaske igakülgse abi eest.
46
KASUTATUD KIRJANDUS
1. Abrams, P.A. 1995. Monotonic or unimodal diversity productivity gradients: What
does competition theory predict? Ecology 76: 2019-2027.
2. Allilender, K., 1997. Soomaa Rahvuspark. Viljandimaa loodus. Viljandi, 4-7.
3. Allilender, K., 2000. Lamminiitude majandamine. Soomaa Rahvuspargi
kaitsekorralduskava 2000 2010. Viljandi. 41 lk.
4. Barot, S. 2004. Mechanisms promoting plant coexistence: can all the proposed
processes be reconciled? Oikos 106: 185-192
5. Benstead, P., Jose P., Joyce, C., Wade, M., 1999. European Wet Grasslands.
Guidelines for management and restoration. RSPB, Sandy.
6. Connell, J.H. 1978. Diversity in tropical rain forests and coral reefs. Science 199:
1302-1309.
7. Diemer, M.W. & Pfadenhauer, J. 1987. Effects of differential defoliation on shoot
growth, density and phytomass of three graminoids in a calcareous fen. Oikos 50: 183-
190.
8. Eriksson, O. 1993. The species-pool hypotesis and plant community diversity. Oikos
68: 371-374.
9. Falinska, K.1991. Plant demography in vegetation succession. Kluver Academic
Publishers, Dortrecht, Boston, London. 1-220.
10. Falinska, K. 1995. Genet disintegration in Filipendula ulmaria: consequences for
population dynamics and vegetation succession. Journal of Ecology 83: 9-21.
11. Grace, J.B. 1999. The factors controlling species density in herbaceous plant
communities: an assessment. Persp. Plant. Ecol. Syst. 2: 1-28
12. Grime, J.P. 1973. Competitive exclusion in herbaceous vegetation. Nature 8London)
242: 344-347.
13. Grime, J.P. 1979. Plant strategies and vegetation processes. John Wiley & Sons,
London.
14. Hallanaro, E.-L., Pylvänäinen, M., Randla, T., 2002. Põhja-Euroopa loodus
Bioloogiline mitmekesisus muutuvas keskkonnas. Nord 2001:15, Põhjamaade
Ministrite Nõukogu. Kopenhaagen, 266-268.
15. Järvet, A., 1994. Soomaa veestik ja jõgede veereziim. ELUS XVII Eesti
loodusuurijate päeva ettekannete kokkuvõtted. Soomaa Rahvuspargi loodus, Tipu, 11.-
12 juuni 1994, 12-18.
47
16. Järvet, A., Karu, M., 1999 Kõik algab jõgedest. Eesti Loodus 10, 410-412.
17. Kasepalu, A., 1991. Mis peremees jätab, selle mets võtab. Tallinn, 5-36.
18. Kink, H., 1996. Soomaa rahvuspark (soo, Pärnu madalik, Sakala kõrgustik). Eesti
kaitsealad geoloogia ja vesi. Tallinn, 111-121.
19. Krall, H.,1979. Rohumaade püsivuse ja tsönootilise suletuse küsimusi. ELUS-i
Aastaraamat. Tartu, 67, 55-64.
20. Krall, H., Pork, K., aug, H., Püss, Õ., Rooma, I, Teras, T. 1980. Eesti NSV loodluslike
rohumaade tüübid ja taimekooslused. Eesti NSV Põllumajandusministeerium, Eesti
NSV TA Zooloogia ja Botaanika Instituut, RPI ,,Eesti Põllumajandusprojekt". Tallinn,
88 lk.
21. Kukk, T., Kull, K., 1997. Puisniidud. Estonia Maritima, 2, 1-249.
22. Kukk, T., 2004. Lamminiidud ehk luhad. Pärandkooslused. Õpik-käsiraamat. Tartu,
191-203.
23. Kumari, E., 1960. Looduskaitse teatmik. Tallinn, 94-96.
24. Laasimer, L., 1965. Eesti NSV taimkate. Tallinn.
25. Leht, M. 1999. Eesti taimede määraja. Eesti Loodusfoto, Tartu
26. Leibak, E., Lutsar, L. 1996. Eesti ranna- ja luhaniidud. Tallinn.
27. Lillema, L., 1958. Eesti NSV Mullastik.Tallinn, 149-181.
28. Loorits, O., 2001. Endis-Eesti elu-olu IV. Lugemispalu karjakasvataja elust. Eesti
Kirjandusmuuseum,Tartu.
29. Loorits, O., 2000. Endis-Eesti elu-olu III. Lugemispalu põllumehe elust. Eesti
Kirjandusmuuseum,Tartu.Luhamaa, H., Ikonen, I., Kukk, T., 2001. Läänemaa
pärandkooslused. Seminatural communities of Läänemaa County, Estonia.
Pärandkooluste Kaitse Ühing, Tartu Turku.
30. Mander, Ü. 1994. Changes of landscape structure in Estonia during the Soviet period.
GeoJournal. Vol 33, No 1, 45-54.
31. Mander, Ü., Palang, H., Tammiksaar, E., 1994. Landscape changes in Estonia during
20th century. Anelysis of Landscape Dynamics Driving Factors Related to Different
Scales. EUROMAB Research Program. Germany, Leipzig-Halle. 73-95.
32. Miller, T.E. 1982. Community diversity and interactions between the size and
frequency of disturbance. Am. Nat. 120: 533-563.
33. Oksanen, J. 1996. Is the humped relationship between species richness and biomass an
artefact dues to plot size? J. Ecol. 84:293-295.
48
34. Paal, J. 1997. Eesti taimkatte kasvukohatüüpide klassifikatsioon. Tartu Ülikooli
Botaanika ja Ökoloogia instituut, Tallinn.
35. Paal, J., Ilomets, M., Fremstad, E., Moen, A., Borset, E., Kuusemets, V., Truus, L.,
Leibak, E., 1999. Eesti märgalade inventeerimine 1997. a. Projekti "Eesti märgalade
kaitse ja majandamise strateegia" aruanne. Eesti Loodusfoto. Tartu, 40-43.
36. Palang, H., Mander, U., Luud, A. 1998. Landscape diversity changes in Estonia.
Landscape and Urban Planning. Vol. 41, No. 3-4, 163-169.
37. Pork, K., 1959. Kesk - Eesti jõgede luhaniitude keskkonnatingimustest. ELUS-i
Aastaraamat. Tartu, 52, 51-70.
38. Pork, K., 1964. Taimkatte genees ja antropogeensed suktsessioonid luhtadel. ELUS-i
Aastaraamat. Tartu, 56, 97-112.
39. Pork, K., 1981. Niidutaimkatte kujunemine, nüüdisaegne seisund ja niitude kasutamise
küsimusi Eesti NSV-s. ELUS-i Aastaraamat. Tartu, 67, 7-37.
40. Pork, K., 1984. Kasari jõe alamjooksu luht geobotaanilise looduskaitseobjektina. Eesti
NSV Riiklike Looduskaitsealade Teaduslikud Tööd IV. Matsalu loodusest. Tallinn,
61-72.
41. Prach, K. 1992. Vegetation, microtopography and water table in the Luznice River
floodplain, South Bohemia, Czechoslovakia. Preslia 64 (4): 357-367.
42. Pärtel, M., Mändla, R., Zobel, M. 1999. Landscape history of a calcareous (alvar)
grassland in Hanila, western Estonia, during the last three hundred years. Landscape
Ecology, 14, 187-196.
43. Pärtel, M. and Zobel, M. 1999. Small-scale plant species richness in calcareous
grasslands determined by the species pool, community age and shoot density. -
Ecography 22: 153-159.
44. Pärtel, M., Zobel, M., Zobel, K. and Van der Maarel, E. 1996. The species pool and its
relation to species richness - evidence from Estonian plant communities. Oikos 75:
111-117.
45. Reintam, L.,1962. Mullateadus. Tallinn.
46. Sarv, T. 1984. Kui palju on Eestis puisniite? Eesti Loodus 6: 360-361
47. Stevens, M.H.H. & Carson, W.P. 1999. Plant density determines species richness
along an experimental fertility gradient. Ecology 80: 455-465.
48. Zobel K., Liira, J. 1997. A scale-independent approach to the richness vs biomass
relationship in ground-layer communities. Oikos 80:325-332.
49
49. Zobel, M. 1992. Plant-species coexistence the role of historical, evolutionary and
ecological factors. Oikos 65: 314-320.
50. Zobel, M. 1997 The relative role of species pools in determining plant species
richness: an alternative explanation of species coexistence? TREE 12: 266-269.
51. Tamm, L. 1967. Süstaga Halliste jõel. Eesti Loodus 6: 383-386.
52. Taylor, D. R., Aarssen, L. W. & Loehle, C. 1990. The species-pool hypothesis and
plant community diversity. Oikos 68: 371-374.
53. Talvi, T., 2001. Pool-looduslikud kooslused. Ökoloogiliste Tehnoloogiate keskus.
54. Tilman, D. 1982. Resource competition and community structure. Princeton
University Press, Princeton.
55. Touzard B., Clément B., Lavorel S. 2002. Successional patterns in an eutrophic
alluvial wetland of western France. Wetlands 22 (1): 111-125.
56. Truus L., Tõnisson, A. 1998. The ecology of floodplain grasslands in Estonia. In:
Joyce CB, Wade PM (eds) European Wet Grasslands: Biodiversity, Management and
Restoration. Wiley, Chichester, 49-60.
57. van der Maarel, E. 1998. Species diversity in plant communities in relation to structure
and dynamics. In: During H. J., Werger, M.J.A. & Willems, J. H. (eds.). Diversity and
pattern in plant communities, SPB Academic Publishing, Hague. 1-14.
58. van Dijk, G. 1991. The status of semi-natural grasslands in Europe. In: The
conservation of lowland dry grassland birds in Europe. Goriup, P. D. (ed.) Newbury:
JNCC Publications, 15-36.
Käsikirjad
59. Karu, M., 2001. Soomaa Rahvuspargi Karuskose ja Lemmjõe lammimetsa
taimekooslused ja mullastik. Magistritöö. Tartu. Käsikiri TÜ Geograafia Instituudis.
60. Meier, E., 2001. Lääne-Saaremaa puisniitude liigirikkuse põhjustest. Tallinn.
Diplomitöö. Käsikiri Tallinna Pedagoogikaülikooli Matemaatika-loodusteaduskonna
Bioloogia õpetoolis.
61. Mikk, H., 1957. Halliste jõe luhtade taimkate (Tipu ja Riisa küla vahelisel alal)
Diplomitöö. Tartu. Käsikiri TÜ Botaanika ja Ökoloogia Instituudis.
50
62. Pullisaar, T., 1959. Materjale Kasari, Halliste ja Navesti luhanitude taimkattest.
Diplomitöö. Tartu. Käsikiri TÜ Botaanika ja Ökoloogia Instituudis.
63. Suurkask, M., 1996. Halliste luha ja puisniitude inventeerimine. Aruanne. Käsikiri
Soomaa Rahvuspargis.
64. Suurkask, M., 1999. Soomaa Rahvuspargi lammirohumaade taimkate ja
suktsessiooniline seisund. Magistritöö. Käsikiri TÜ Botaanika ja Ökoloogia
Instituudis.
51
Üleujutuse mõju taimkattele
Sarnased õppematerjalid
![Eesti rahvuspargid]()
13
doc
Eesti rahvuspargid
rabad ja sood.
Samamoodi nagu Karula loodus on väga mitmekesine, on kirju ka selle Eestimaa osakese
minevik. Sajandite vältel on vana kihelkonda mitmel viisil jaotatud, võimud vahetunud,
inimeste kombed muutunud, rahvastik sõdades vähenenud ja rahu ajal jälle kasvanud.
Karula rahvuspark paikneb enam-vähem ajaloolise Karula kihelkonna piirides, hõlmates
kihelkonna idaosa. Ajaloo vältel toimunud administratiivsete piiride muutustest hoolimata on
kihelkond püsinud tervikuna - inimesi seovad võru keel, vanadest aegadest pärit igapäevaelu
kombed ja harjumused, läbi sajandite püsinud sugulussidemed ning kõrgustiku loodusest
tulenev elustiil. Säilinud on sajandivanused taluhooned ja põllupaigad, suust-suhu liiguvad
iidsed rahvajutud, teatakse muinasaegsete kivikalmete asukohti, räägitakse metsavendadest
ning hoitakse korras esivanemate haudu Lüllemäel ja Kaikamäel.
Karula inimene paikne ja traditsioone austav
![Biotoopid]()
54
docx
Biotoopid
mineraalaineid vähem.
• Alanemisfaas – seisev vesi jõe kaldavalli taga, vetikate arenemine.
Setted
• on luhtade mullaviljakuse ja produktiivsuse aluseks.
• on sageli lubjarikkad, sisaldavad rohkelt fosforit.
• maksimaalne settekiht 7 cm, enamasti 2–25 mm.
• sadeneb ajas ja ruumis erinevalt.
Esinduslikemate Eesti luhamassiivide iseloomustus.
Soomaa luhad
• Keskseks voolusooneks on Navestisse suubuv Halliste jõgi millesse suubub
omakorda Raudna jõgi, millel on vaja vastu võtta Lemmjõe ja Kõpu jõe
veed.
• Üleujutatud ala pindalalt suurim kogu Eestis, nõrga üleujutuse korral 3000-
4000 ha, keskmisel aastal 5000 hektarit, maksimaalse veeseisu korral
umbes 21000 hektarit (u. Võrtsjärve pindala).
• Suurem osa Soomaa luhaniite on tekkinud lammimetsadest. Lagedat
lammi ilmestavad üksikud puud ja põõsad andes talle kohati hõreda
![Pärandkooslused eksam]()
16
doc
Pärandkooslused eksam
Taimekooslus - koos kasvavate taimede kogum.
Puisniit - puude ja põõsastega heinamaa ehk regulaarselt niidetava rohustuga hõre puistu.
Fluktuatsioon - koosluse/ökosüsteemi muutumine mõne aasta kuni kümnete aastate jooksul.
Muutub liikide arvukus, osakaal koosluses, kuid koosluse üldilme jääb samaks.
Suktsessioon – koosluse/ökosüsteemi muutumine sadade kuni tuhandate aastate jooksul.
PKÜ – Pärandkoosluste kaitse ühing
ELF – Eestimaa looduse fond
Millal alustati niitude taimkatte uurimist Eestis ning millal ja miks hakati tähelepanu
pöörama rohumaade kaitsele?
Geobotaaniline uurimine algas 19. sajandi II poolel. Enne II maailmasõda ilmus palju
rohumaaviljeluse käsiraamatuid, uuriti rohumaade saagikuse suurendamise võimalusi, niitude
kultuuristamist. Niitude kaitsega ei tegeletud. 1950ndatel uuriti niitude saagikuse tõstmist
pealtparandamise abil. 1960ndatest tegeletakse poollooduslike rohumaade kaitsega: Pork,
Krall, Kull jt.
Rohumaade kasvukohatüübid
1
![Pärandkoosluste eksam]()
28
docx
Pärandkoosluste eksam
kooslust, kus inimtegevusega (väetamine, heinaseemne täiendav külv jms.) on tõstetud
rohumaa kasutusväärtust, kuid seejuures säilib rohukamaras enamik varem kasvanud
taimedest.
Karjamaa - rohumaa, kus karjatatakse kariloomi. Puudega karjamaid nimetatakse
puiskarjamaadeks. Eristatakse looduslikke karjamaid, mis on pärandkooslused, ning
kultuurkarjamaid, mis on külvatud rohustuga.
Niit e heinamaa - ökosüsteem, milles taimkatte moodustavad peamiselt tihedalt
kasvavate mitmeaastaste mesofüütide kooslused, mille mullas toimub kamardumine ja
mida võidakse regulaarselt niita.
Primaarsed niidud on rohumaad, mis on kujunenud ilma inimese olulise
osaluseta. Primaarseiks niitudeks võivad Eestis olla mõned lamminiidud (kus
üleujutuse ja jää mõjul ei kasva puid) ja mõned rannaniidud (eelkõige maatõusu
alal, mis veel pole metsastunud).
![Eesti biotoobid]()
48
docx
Eesti biotoobid
• Lamminiidud asuvad jõgede, harvemini järvede madalatel tasastel kaldaaladel (lammidel).
• Enamus Eesti lamminiite on sekundaarse inimtekkelise päritoluga, eriti jõgede keskjooksudel, kus
levisid varem lammimetsad.
3.2. Esinduslikemate Eesti luhamassiivide iseloomustus. – soomaa- enamjaolt tekkinud
lammimetsadest, ilmestavad üksikud puud ja põõsad. Säilinud luhaniite 1000ha. Üleujutatud ala
pindalalt eesti suurim. , kasari – suurus u 4000ha. Luha viljakus märgatavaltkõrgem kui enamikul teiste
eesti jõgede luhtadel. Muld väga viljakas.
Soomaa luhad: • Suurem osa Soomaa luhaniite on tekkinud lammimetsadest. Lagedat lammi ilmestavad
üksikud puud ja põõsad andes talle kohati hõreda puisniidu ilme. • Säilinud luhaniite on Soomaal umbes
1000ha, neist hooldatavad vaid ca 600. • Keskseks voolusooneks on Navestisse suubuv Halliste jõgi
millesse suubub omakorda Raudna jõgi, millel on vaja vastu võtta Lemmjõe ja Kõpu jõe veed
![Pärandkoosluste eksamiks]()
11
docx
Pärandkoosluste eksamiks
Üldmõisted
Pärandkooslused ( pool-looduslikud kooslused) on inimese ümberkujundatud looduslikud kooslused eelkõige puisniidud, loopealsed,
lamminiidud, rannaniidud, aga ka teised karja- ja heinamaad, mis sellisena püsivad mõõduka inimmõju (niitmine, karjatamine) tingimustes.
Niitmine, valikraie, ekstensiivne karjatamine, kulupõletus on koosluse muutmise looduslähedased viisid, mis ei vii valdava osa liikide
väljalangemisele nagu kamara ümberkündmine, mulla teisaldamine, tugev väetamine või mürgitamine. Inimmõju lõppemisel muutuvad
pärandkooslused loodusliku suktsessiooni käigus looduslikuks koosluseks (enamasti metsaks).
Pärandkooslused klassifitseeritakse: esimeseks on aruniidud, mis on turvastumata ja üleujutuseta muldadel ning jaotuvad looniitudeks e.
alvariteks e. loopealseteks(s.h. loopuiskarjamaad), pärisaruniitudeks (s.h. pärisarupuisniidud ja karjamaad), nõmmeniitudeks ja
paluniitudeks, Teiseks on lamminiidud e. luhad (s.h. lammipuisniidud), kolmanda
![Eesti looduskaitse]()
26
pdf
Eesti looduskaitse
4) olulisemate kokkuvõtvate raamatute ilmumine. 1929 töötas komisjon (Andres Mathiesen, Edmund
rahvuspark Ameerika Ühendriikides (1872), Saksamaa Spohr, Artur Luha, Gustav Vilberg (Vilbaste)) 1947 moodustati 7 jahikeeluala: Vigala, Matsalu,
linnukaitseseadus (1888) Nimetamata on jäänud nüüdisajal aktiivselt välja esimese looduskaitseseaduse projekti. Vilsandi, KursiKärevere, Tudulinna, Kuressaare
ja 9 rahvusparki Rootsis (1909)
Keskkonna ja loodusõpetus
![Referaat Pärandkooslused]()
9
docx
Referaat Pärandkooslused
nendel aladel loomulik. (Sammul, M. jt 2003)
Lamminiidud
Lamminiidud ehk luhad on jõgede kallastel asuvad niidud, mida suurvesi perioodiliselt üle
ujutab. Luhad on tekkinud aladele, kust inimene on lammimetsa maha võtnud ning karjatama
ja niitma hakanud. Vähemal määral leidub ka primaarseid lamminiite jõeloogetes ja jõgede
deltaaladel.
Lamminiitude taimestikku iseloomustavad erinevad jõega paralleelselt kulgevad ning luha
kõrgusest ja üleujutuste ulatusest sõltuvad vööndid. Jõest kaugemal, kuhu tulvaveed jõuavad
harva, domineerib kuiva lamminiidu kasvukohatüüp, mis on üsna liigirikas ja kus on
enamuses niidutaimed. Mida jõele lähemale, seda niiskemaks muutub maa ja kõrgemaks
taimestik. Jõele lähemal niisketes tingimustes domineerivad suuremad tarnad ning maa võib
�olla isegi soostunud. Ka lamminiitudel on vähese majandamise tõttu hakanud vohama
roostikud.
Kommentaarid (0)





































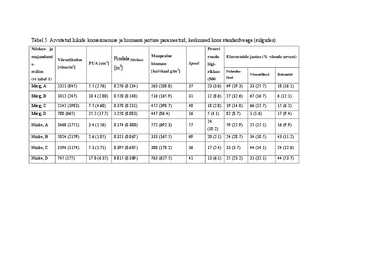
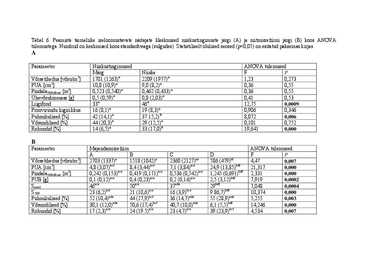































Kõik kommentaarid