mikrobioloogia (0)
· Inimese mikroobikooslus
Inimese normaalne mikrobioota: mõisted, nende erinevused.
mikrobioota - mikroobikooslus ühes kehaosas
mikrobioom - kõik mikroobid, nende komponendid ja aktiivsused kokku
Normaalse mikrobioota funktsioonid inimese jaoks ja faktorid, millest selle
koostis sõltub.
– Mikrobioomi funktsioonid: immuunsüsteemi “välajõpe”, kiudainete lagundamine,
vitamiinide süntees sooles, rasvhapete moodustumine sooles, sooleepiteeli
tugevdamine, kaitse patogeenide eest, ravimite töötlemine, meeleolu mõjutamine.
Mikrobioota koostis sõltub vanusest, sünni viisist, toidust, elukeskkonnast, tervislikust
seisundist, ravimite kasutamisest jne.
Kes on gnotobiondid?
– organismid, kelle mikrobioota on määratletud. Nad võivad olla steriilsed ehk
mikroobivabad loomad, aga ka loomad, keda on ekperimentaalselt nakatatud mingite
kindlate mikroobidega. Imetajatel saadakse mikroobivabu loomi keisrilõikega, sest
vastsündinu on emaihus praktiliselt steriilne. Mikroobid saadakse sünniteedest ja
juurde inimeste nahalt, emapiimast ja väliskeskkonnast.
Mikroobivabade loomade eripärad.
– elavad steriilsetes tingimustes kauem kui tavalised liigikaaslased, neil on õhem soole
limaskest ja toitained imenduvad kiiremini, vitamiini vajadus suurem, väljaheidete
kogus väiksem, immuunsüsteem on nõrgem, mikroobid, mis ei põhjusta haigestumist
tavalistel loomadel on patogeensed mikroobivabadele loomadele. Nende veres on
kordades vähem metaboliite.
Inimese erinevate kehapiirkondade (naha, mao, suuõõne, soolestiku)
mikrobioota tunnused ja selle põhirollid.
– Ülemistes kuseteedes pole mikroobe. Naha mikrobioota on harjunud kuiva ning
happelise pinnaga, halotolerantsed, toodavad naharasu lipiididest orgaanilisi happeid,
mis on bakteritsiitsed.
Suuõõnes on palju mikroobe, sülg sisaldab lüsosüümi, mis lüüsib baktereid, bakterid
võivad põhjustada hambakaariest ja hambakattu, igemetaskutes elavad anaeroobsed
bakterid on võimelised tungima ka epiteelirakkudesse ja sügavamatesse kudedesse,
põhjustades nende kahjustusi. Tekib põletikuline protsess hammast ümbritsevates
kudedes: igemed veritsevad, hambad hakkavad loksuma jmt. Mao happeline keskkond
pärsib bakterite ellujäämist maos. Neist kõige arvukam on Helicobacter pylori, kes
osaleb ka maohaavade ja maovähi tekitamises. Ta toodab ureaasi ja tekitab enda
ümber ammoniaagipilve, mis neutraliseerib hapet. Kuna helikobakter aitab mao
ülihappelisust vältida, ei pruugi ta alati kahjulik olla. Soolestiku bakterid on enamasti
anaeroobid või fakultatiivsed anaeroobid, nad aitavad lagundada suhkruid, oituvad
ainetest, mis ülemistes seedekulgla osades seedimata ja imendumata jäävad
(kiudained) ning ka irdunud sooleepiteelist ja soolelima suhkrutest ja valkudest
(mutsiinid). Mutsiinid on glükosüülitud valgud. Soolelima valgud ehk mutsiinid on
soolebakteritele toiduks.
Miks on oluline Malassezia?
– kandseente hulka kuuluv pärm, mis põhjustab kõõma ja nahaekseeme. Võib
moodustada filamente ja pseudomütseeli. Malassezia on lipofiilne, ta kuulub
soojavereliste loomade naha normaalsesse mikrobiootasse, kuid vohades võib
põhjustada haigusi. Seene vohamist soodustab naha rasunäärmete eritis, mis sisaldab
talle vajalikke lipiide. Osa Malassezia liike vajab obligaatselt lipiide.
Biokeemilised protsessid jämesooles, mida viivad läbi soolebakterid.
– Lagundab suhkruid, metaboliseerivad sapphappeid, lagundavad ja kääritavad
kiudaineid lühikese ahelaga rasvhapeteks (atsetaat, propionaat, butüraat) ja
piimhappeks. Propionaadist sünteesitakse maksas glükogeeni. Atsetaadist
sünteesitakse maksas ja rasvarakkudes rasvu (lipogenees). Atsetaat mõjutab ka
histoonide atsetüülimist ja sellekaudu geenide ekspressiooni (epigeneetika). Butüraat
on põhiliseks energeetiliseks aineks sooleepiteeli rakkudele –
kolonotsüütidele. Hoiab epiteeli intaktsena ja kaitseb soolevähi eest. Need kolm
rasvhapet on põhilised käärimisel moodustuvad rasvhapped ka mäletseja looma
vatsas (rumen). Vatsas seedivad taimseid kiudaineid anaeroobsed bakterid.
Sünteesivad sooles vitamiine, serotoniini, nende sünteesitud metaboliidid mõjutavad
inimese närvitalitust, käitumist ja emotsioone.
Käärimisel moodustuvad rasvhapped ja gaasid.
– Propionaat, atsetaat, butüraat tekivad kääritamisel koos süsihappegaasi, metaani ja
veega.
Butüraat kui oluline metaboliit jämesooles, kuidas tekib ja milline on selle
roll.
– on põhiliseks energeetiliseks aineks kolonotsüütidele. Butüraadi oksüdatsioon
hapnikuga epiteelirakkudes hoiab soole epiteelipoolse osa anaeroobsena. Anaeroobne
keskkond sobib soolebakteritele. Mõned neist on väga hapnikukartlikud. Kui
epiteelirakud on hea tervise juures, on ka sooleepiteel terve ega leki. Terve
sooleepiteel on ka paremini kaitstud soolevähi eest. Ka soolelima kihi paksus on
tervise seisukohalt oluline. Kui limakiht on õhuke, on suurem oht lekkeks soolest või
infektsiooniks.
Millised kiudained ja miks toimivad jämesoolebakterite põhitoiduna?
–Nad toituvad ainetest, mis ülemistes seedekulgla osades seedimata ja imendumata
jäävad, milleks on kiudained. Soolebakterid on evolutsioneerunud koos inimesega ja
kohanenud nende ainete kasutamiseks, mida jämesooles leidub.
Prebiootikumid ja probiootikumid, nende definitsioonid ja mõned näited.
– Probiootikumid (probiootilised bakterid) on tervete inimeste (ja loomade) soolestikust
isoleeritud elusad mittepatogeensed baktereid, mille (suukaudne) manustamine mõjub
soodsalt inimese (looma) tervisele. Probiootikumidega kasutatakse peamiselt
bifidobaktereid ja laktobatsille. Prebiootikumid on bakterid, mis mõjuvad soodsalt
organismi tervistning nad toimivad bakterite vahendusel, kellele nad on toiduks.
Näiteks insuliin, oligufruktoos, galaktooligosahhariidid. Prebiootikumid - probiootikume
stimuleerivad ained.
Igemehaiguste mikrobioloogilised põhjused.
– Igemetaskutes elavad anaeroobsed bakterid on võimelised tungima ka
epiteelirakkudesse ja sügavamatesse kudedesse, põhjustades nende kahjustusi. Tekib
põletikuline protsess hammast ümbritsevates kudedes: igemed veritsevad, hambad
hakkavad loksuma jmt. Sageli on parodontoosiga inimestel suus vähe
piimhapebaktereid ja kaariest, st välja kukuvad terved hambad. Olulisimad
parodontoosi põhjustajad on gramnegatiivsed anaeroobsed bakterid Aggregatibacter
actinomycetemcomitans, Porphyromonas gingivalis, Prevotella intermedia,
Fusobacterium nucleatum, Capnocytophaga liigid jt. Aggregatibacter
actinomycetemcomitans on gramnegatiivne pulkbakter, keda sageli leitakse
agressiivse paradontoosi korral. Moodustab biofilmi. Palju erinevaid
virulentsusfaktoreid. Kahjustab ka leukotsüüte. Bakterist raske lahti saada. On
katsetatud ka töötlemist bdellovibrioonidega, kes lüüsivad elusaid rakke. Probleemiks
on see, et parodontoosi põhjustajad on hapnikukartlikud bakterid ja bdellovibrioonid
vajavad hapnikku.
· Antibiootikumid
Milleks kasutatakse antibiootikume ja sulfanüülamiide?
– Antibiootikumid (ja sulfaniilamiidid) on ained, mida kasutatakse bakter- ja
seenhaiguste ravil.
Antibiootikumi definitsioon.
– aine, mida kasutatakse bakteri- ja seenhaiguste ravil.
Antibiootikumide produtsendid.
– keemiline süntees (sulfaniilamiid, isoniasiid), mikroorganismide sünteesitud,
keemiliselt modifitseeritud (poolsünteetilised penitsilliinid). Põhilised antibiootikumide
sünteesijad on hallitusseened ning aktinobakterid. Mõningaid antibiootikume toodavad
ka batsillid, müksobakterid ja lüsobakterid. Mikroorganismid sünteesivad
antibiootikume aktiivse kasvu lõppedes, üleminekul statsionaarsesse kasvufaasi.
Seega võib AB käsitleda sekundaarsete metaboliitidena.
Antibiootikumide põhigrupid, nende põhilised märklauad ja
toimemehhanismid rakus.
– Kui AB põhjustab bakteriraku surma, siis on tegu bakteritsiidse AB-ga. Kui aga AB
ainult surub alla bakteri kasvu, siis nimetatakse AB bakteriostaatiliseks. AB, mis toimib
paljudele erinevatele bakteritele, on laia toimespektriga. AB, mis toimib ainult
vähestele bakteritele, on kitsa toimespektriga. Erütromütsiin, tetratsükliin ja
sulfaniilamiidid on laia toimespektriga, isoniasiid mõjub aga ainult mükobakteritele,
sest tema märklauaks on mükoolhapete sünteesi rada. Sulfaniilamiidi märklauaks on
foolhappe sünteesirada. Foolhappe sünteesirada on vajalik ühesüsinikulisteks
ülekanneteks biosünteesiradades. Sulfanüülamiidid on PABA struktuuranaloogid. Kui
sünteesis kasutatakse PABA asemel sulfanüülamiidi, siis foolhappe sünteesirada
blokeerub ja bakteri kasv seiskub.
Struktuuri järgi saab jagada antibiootikumid: aminoglükosiidsed AB-d (märklauaks
valgusüntees), tetratsükliinid (märklauaks valgusüntees), makroliidid, polüpeptiidsed
AB-d (takistab peptidoglükaani sünteesi), kinoloonid, polüeensed AB-d, glükopeptiidid,
beeta-laktaamsed AB-d (sisaldavad beeta-laktaamituuma), peptiidsed AB-d.
Anitobiootikumid jagatakse toimemehhanismide järgi rühmadeks: Rakukesta sünteesi
takistavad AB-d, Valgusünteesi takistavad AB-d, Membraanidele toimivad AB-d,
Metaboolsed antagonistid (normaalsete metaboliitide struktuuranaloogid), Rakuskeleti
valkudele (FtsZ jt) mõjuvad AB-d (uus märklaud), Patogeenide virulentsusfaktorite
sekretsioonile (Tüüp III sekretsioonisüsteem) mõjuvad AB-d (uus spetsiifiline
märklaud). Seda süsteemi kasutavad näiteks katku, düsenteeria ja tüüfuse tekitajad,
Siderofooride sünteesi pärssimine (bakteri jätmine rauanälga), Kvoorumi tunnetamist
häirivad AB (potentsiaalne märklaud), Adhesiooni takistamine.
Kuidas võib tekkida resistentsus, kuidas seda vältida ja vähendada?
– Võib tekkida kui antibiootikum ei pääse rakku, pääseb rakku, aga pumbatakse kohe
aktiivselt rakust välja, antibiootikumi märklaud puudub, märklaua hulk suureneb, kui
antibiootikum lagundatakse või modifitseeritakse, kui rakus tekib alternatiivne
metaboolne rada, mida antibiootikum ei suuda mõjutada. penitsiliin toimib ainult
noortele jagunevatele rakkudele mitte vanadele. Lisaks β-laktamaasidele on
resistentsuse tekke põhjuseks: AB ei suuda tungida rakku (välismembraani poriinide
geenides on toimunud mutatsioonid), Transpeptidaas muteerub ega suuda enam AB
siduda, Mõnel bakteril (Pseudomonas aeruginosa) on kirjeldatud ka β-laktaame
väljapumpavaid transportereid – pumpasid. Bakterite ribosoomid on teistsuguse
ehitusega kui eukarüootide tsütoplasmaatilised ribosoomid. Seeõttu valgusüntees
bakterites selektiivne märklaud AB-dele. Resistentsuse teke aminoglükosiidsetele AB-
dele: ribosoomi seostumissaitide metüleerimine, AB modifitseerimine (adenüülimine,
fosforüülimine, atsetüleerimine). Resistentsus makroliitidele tekib: märklaua
muteerumisega, AB väljapumpamisega, märklaua modifitseerimisega (ribosomaalse
RNA metüleerimine), AB modifitseerimisega (fosforüülimine, glükosüleerimine), AB
lagundamisega. AB tootvad bakterid väldivad enesetappu sellega, et neil puudub AB
märklaud (näiteks penitsillini sünteesivatel hallitusseentel ei ole peptidoglükaani), nad
modifitseerivad endal selle märklaua, millele nende toodetav AB seostub (ribosoomi
RNA-de metüleerimine), märklaud, millele AB toimib, on produtsendis muteerunud ja
seetõttu AB-le tundetu, kohe peale AB sünteesimist pumpavad selle kiiresti välja,
rakust väljasuunatud AB seostub väliskeskkonnas näiteks mullaosakestele, millega
väheneb tema tagasitransportimise võimalus rakku.
Mikroobide resistentsust antibiootikumidele saab piirata sellega, et tarvitada AB
määratud koguses (piisav doos) ja vajalik arv päevi (läbida terve kuur), kasutada
kombineeritud AB-sid (nt sulfaniilamiid + trimeoprim), piirata (keelata) AB-de
kasutamist loomade kasvustimulaatorina. EU-s seda enam ei lubata. Aga kasutatakse
loomade ravimisel ja epideemiate ennetamisel. Veterinaarias soovitatakse kasutada
neid ravimeid, mida inimeste ravimisel ei kasutata. Vahetult enne loomade tapmist ei
tohiks AB-sid kasutada, piirata AB kasutamist taimekasvatuses (tetratsükliin,
streptomütsiin).
Milline on resistentsusgeenide päritolu ja kuidas need mikroobide hulgas
levivad?
– Päritolu: olid olemas juba enne AB-de massilist kasutuselevõttu meditsiinis, on
ilmselt pärit AB-sid tootvatest mikroobidest, kellest on levinud teistele bakteritele
horisontaalse geeniülekandega ja plasmiididega, penitsillinaas on kujunenud
transpeptidaasi geeni baasil: geen duplitseerus ja üks geenikoopia evolutsioneerus
penitsillinaasi geeniks. Penitsillinaas ja transpeptidaas on struktuurselt sarnased.
Levivad transformatsiooniga, konjugatsiooniga (plasmiidide ülekandumisel),
transduktsiooniga (bakteriofaagide abil). Tihti paiknevad resistentsusgeenid kas
plasmiididel või transposoonidel (mobiilsed elemendid).
Antibiootikumide märklauad bakteritel, mida saab mikroobidega võitlemisel
kasutada.
– Rakukesta sünteesi takistavad AB-d, Valgusünteesi takistavad AB-d, Membraanidele
toimivad AB-d, Metaboolsed antagonistid (normaalsete metaboliitide
struktuuranaloogid), Rakuskeleti valkudele (FtsZ jt) mõjuvad AB-d (uus märklaud),
Patogeenide virulentsusfaktorite sekretsioonile (Tüüp III sekretsioonisüsteem) mõjuvad
AB-d (uus spetsiifiline märklaud). Seda süsteemi kasutavad näiteks katku, düsenteeria
ja tüüfuse tekitajad, Siderofooride sünteesi pärssimine (bakteri jätmine rauanälga),
Kvoorumi tunnetamist häirivad AB (potentsiaalne märklaud), Adhesiooni takistamine.
Uued võimalused on nanoosakesed (biokile tekke pärssimine), hulgatunnetus
(bakterite suhtlemise ja biofilmi tekke pärssimine), siderofoorid (ensüümidelt
kofaktorite äta võtmine), aluselised peptiidid (membraanide lõhkumine), aluselised
nanoosakesed (membraanide lõhkumine), vaktsiinid (bakterite antigeenide sidumine),
metalliioonid (membraani lõhkumine), immuunsuse komponendid (antikehade ja AB-
de konjugaadid), bakteriofaagid (membraanide lüüsimine), suhkrute analoogid
(suhkruid metaboliseerivate ensüümide häirimine), suhkrud (adhensioomi
blokeerimine), pilitsiidid 8piilide moodustumise inhibeerimine).
· Mikroorganismide toitumine ja ATP süntees
Milline on mikroobide eripinna ja kuju seos toitumisega?
– Suure eripinnaga bakteritel on suurem toitumispind. Sama difusioonialaga raku
ümber on näidatud pulkbakter ja spirill. Spirill saab siiski aineid paremini kätte, sest
tema rakumembraanil on rohkem pinda mahutamaks sinna näiteks transportereid. Kui
toitu on vähe, siis rakud pikenevad ehk filamenteeruvad. Sellega tekib aktiivset
rakupinda juurde. Paraneb ka pinnale kleepumine.
Millised probleemid võivad toitumisega väga suurtel bakteritel tekkida?
– Lihtsad üherakulised organismid (bakterid) ei saaks olla väga suured, sest suurena
nad ei toimiks: nad ei suudaks rakku varustada toitainetega ja aineid raku piires
piisava kiirusega edasi toimetada.
Millised on võimalused eripinna suurendamiseks?
–
Mille poolest on eriline Pelagibacter ubique?
– Väike ja väga suure eripinnaga kõverdunud merebakter. Moodustab enamuse
heterotroofsete merebakterite biomassist. Tema genoom on AT-rikas, mis sisaldavad
summaarselt vähem lämmastikku.
Mikro-ja makroelementide olulisus ning väävli, fosfori ja lämmastiku roll
mikroobide toitumises.
– Väävel kuulub Cys, Met, tiamiini, CoA, biotiini ja lipohappesse. Söötmetesse
pannakse sulfaati ammooniumsulfaadina, mis on nii S- kui ka N-allikaks. Väävlit
saadakse sulfaadist, mis transporditakse rakku ATP energia arvel. Et redutseerida
sulfaati, tuleb see esmalt aktiveerida ATP arvel fosfoadenosiinfosfosulfaadiks (PAPS) ja
seejärel toimub redutseerimine sulfiidini. Vajatakse ka NADPH-d
Fosfor kuulub nukleiinhapetesse ja fosfolipiididesse. Mikroobid saavad kasutada nii
orgaanilisi kui ka anorgaanilisi fosforiühendeid. Fosfori põhikandja rakus on ATP. See
moodustub kas substraatsel või membraansel fosforüülimisel. Fosfaadid on ka head
puhverdajad – fosfaatpuhver. Lisatakse mikroobide kasvusöötmetesse fosfaate pH
puhverdajatena. Osa fosfaati talletatakse ka varuaine polüfosfaadina (volutiin).
Polüfosfaat on kasutatav nii ATP sünteesiks kui ka fosforüüli doonorina
kinaasireaktsioonides.
Lämmastik on rakuaine põhikomponent. Kõige enam on baktereid, kes on suutelised
kasutama orgaanilisi N-ühendeid (valke, aminohappeid, N-aluseid). Need on
heterotroofsed bakterid ehk saproobid. Kasvavad hästi puljongsöötmetel (LB sööde jt).
Nad kasutavad orgaanilisi N-ühendeid nii energia hankimiseks kui ka biosünteesiks.
Osa mikroobe suudab lagundada ka uureat (ureaas) ja kasutada seda N-allikana. Raku
sees moodustub NH3 nitraatide redutseerimisel, õhulämmastiku redutseerimisel ja
orgaanilistest ühenditest (näiteks valkudest) nende hüdrolüüsil. Paljud bakterid saavad
N-allikana kasutada õhulämmastikku N2 Redutseerivad selle ammoniaagiks. Nitraadi
redutseerimises osalevad tsütoplasmaatilised ensüümid nitraadi ja nitriti reduktaas.
Mõlemad kasutavad redutseerijana NADPH2. Kuna nitraadi kasutamine on rakule
energeetiliselt kulukam, kui ammooniumlämmastiku kasutamine, siis reguleeritakse
nitraadi kasutamist ammooniumirepressiooniga: kui keskkonnas on
ammoniumlämmastik olemas, siis ei sünteesita nitraadi transporterit ja nitraadi
redutseerimiseks vajalikke valke või inaktiveeritakse olemasolev nitraadi permeaas.
Miks on arseen inimesele mürgine?
– See on fosforiga sarnane ning valgu kinaasid võivad valgu fosforüülimise asemel
panna valgu koostisesse arseeni.
Mis on kasvufaktorid, prototroofid ja auksotroofid?
– Kasvufaktorid on orgaanilised ained, mida mikroorganismid ei suuda ise sünteesida
ja vajavad neid tavaliselt mikrokogustes. Kasvufaktorit sünteesivat mikroorganismi
nimetatakse prototroofiks, seda, kes ise ei suuda sünteesida, aga auksotroofiks. Kui on
teada, et mikroob on auksotroofne, aga pole teada, milliste kasvufaktorite suhtes, siis
lisatakse söötmetele pärmiekstrakti, peptooni, kasaminohappeid (kaseiini hüdrolüüsil
saadud aminohapete segu) või trüptooni.
Põhilised toitumistüübid, kust saadakse süsinik, elektronid ja energia. ATP ja
prootongradient energiavormidena, kuidas need saadakse.
– Energia saadakse kas valgusest (fototroofid) või keemilistest ainetest (kemotroofid).
Oksüdeeriv aine (elektroni doonor) on kas anorgaaniline aine (litotroofid) või
orgaaniline aine (organotroofid). Ka fototroofid vajavad fotosünteesil elektroni
doonoreid CO2 redutseerimiseks. Nendeks võivad olla näiteks vesi (tsüanobakterid ja
taimed), H2S ja S ja mõned orgaanilised ained (rohelised ja punased fotosünteesivad
bakterid). Süsiniku allikaks on kas CO2 (autotroofid) või orgaanilised ained
(heterotroofid). Prootongradiendi arvel saab sünteesida ATP-d ja teha tööd: keemilised
ained ja anorgaanilised -> elektronide transport -> prootongradient -> ATP/viburite
tööle panek, ainete aktiivne transport rakku. Valgus -> elektronide transport ->
prootongradient -> ATP/viburite tööle panek, ainete aktiivne transport rakku.
Substraatne ja membraanne fosforüülimine, millistes protsessides vastavad
fosforüülimised esinevad.
– Substraatsel fosforüülimisel moodustub ATP fosforüülrühma ülekandmisel ADP-le
mõnelt makroergiliselt katabolismi vaheproduktilt (energiarikkalt substraadilt, nt PEP-
ilt). Substraatne fosforüülimine on praktiliselt ainsaks ATP sünteesi võimaluseks
kääritajatele! Substraatses fosforüülimises osalevad ensüümid on lahustuvad, asuvad
tsütoplasmas. Membraanne fosforüülimine: prootonpump ja ATP süntaas.
Membraanne fosforüülimine toimub membraanidel (mitokondri sisemembraanis
eukarüootidel ja rakumembraanis prokarüootidel). Membraanis paiknevad vaheldumisi
vesiniku ja elektronide ülekandjad. H ülekandjad (flaviinid ja kinoonid) võtavad vastu
nii elektroni kui prootoni, kuid annavad edasi vaid elektroni. Elektronid ei saa
bioloogilistes süsteemides vabalt (mitteseotult) olla, vaid nad liiguvad doonorilt (nt
NADH) aktseptorile. Lõpuks jõuavad nad elektonide lõppaktseptorile (aeroobsel
hingamisel hapnikule). Elektronide liikumine toob kaasa prootonite suunamise läbi
membraani välja. Moodustub prootongradient. Prootonid ei saa vabalt läbi membraani
tagasi difundeeruda – seega moodustunud gradient püsib. Prootonid saavad rakku
tagasi liikuda piki prootongradienti läbi membraanis paikneva ATP süntaasi kanali.
Sellega kaasneb ATP süntees – membraanne fosforüülimine.
Elektrontransportahela komponendid.
–
Kuidas luuakse prootongradient membraanile?
– Elektronid ei saa olla vabalt vaid liiguvad toonorilt aktseptorile ning hingamisahela
lõpus toimub suunatud elektronide liikumine läbi membraani ja see toob kaasa
prootongradiendi moodustumise.
Kuidas toimivad tsüaniid, dinitrofenool ja CO hingamisahela mürkidena?
– Tsüaniid, CO ja asiid (NaN3) seostuvad raua aatomiga tsütokroomi oksüdaasis ja
takistavad elektronide ülekannet hapnikule. Antimütsiin A (antibiootikum) takistab
elektroni ülekannet tsütokroom b-lt tsütokroom c-le.
· Bioohutus
Milles seisneb töökeskkonna riskianalüüs?
– Mõeldakse läbi, millised on potentsiaalsed ohud selles laboris, seda eksperimenti
tehes ning kaalutakse, kas eksperimendi võimalik tulem on seda väärt.
Milliste ohtudega tudeng või töötaja mikrobioloogia laboris kokku võib
puutuda?
– Bioloogiliste ohtudega (mikroobid, patogeenid jne), keemiliste ohtudega (ristsaastus,
reagendid, happed jne), füüsikaliste ohtudega (elektriseadmed, kiirgused jne),
füsioloogilised ja psühhosotsiaalsed ohutegurid (kollegide pahatahtlikkus, kollegide
segamine, üleväsimus jne)
Kuidas neid ohte vältida?
– Tuleb alati käsitled tundmatuid mikroobe kui kõrgeima ohurühma mikroobe, kanda
kaitseriietust ja kasutada steriilseid töövahendeid laminaarkapi all. Mitte
korduvkasutada kindaid. Töötada puhanud olekus ja ratsionaalselt mõeldes. Arvestada
alati keemiliste ainete toksilisusega ja kasutada seadmeid vastavalt juhistele.
Mis vahe on ohul ja riskil?
– Oht on teoreetiline olukord, kus ohutegur kedagi/midagi ähvardab. Risk on selle ohu
realiseerumise tõenäosuse ja mõju tulem.
Mis on ja milleks on tarvis head laboritava?
– Kujundab töökultuuri, mis vähendab bioloogilisest ohuallikast tulenevat riski
töötajale, töökeskkonnale, eksperimendi kvaliteedile ja looduskeskkonnale. GMPP
puhul ei tehta üldiselt vahet kasutatavatel mikroobide ohurühmadel ega laborite
ohutusklassidel – tuleks rakendada alati. See tagab ohutuse laboris.
· Heksooside katabolism
Orgaaniliste ainete aeroobne ja anaeroobne oksüdatsioon.
– Mõlemal juhul toimub glükolüüs, mille tulemusena tekib püruvaat, mis võib
aeroobsetes tingimustes oksüdeeruda edasi tsitraaditsüklis või anaeroobsetes
tingimustes muutuda käärimisproduktideks (fakultatiivsed anaeroobid). Aeroobsel
hingamisel tekib AcCoA püruvaadi dehüdrogenaasikompleksi vahendusel.
Anaeroobidel toimub püruvaadi oksüdeerimine ferredoksiiniga (Fd). Neil puudub NAD-
seoseline püruvaadi dehüdrogenaasikompleks. Fd on mitteheemne rauda sisaldav
valk. Fd- seoseline oksüdeerimisreaktsioon esineb näiteks klostriididel
käärimisradades (võihapekäärimine). Ac-CoA-st moodustuvad käärimisproduktid, nt
atsetaat.
Tsitraaditsükkel, selle funktsioonid.
– Ensüümide katalüüsitud biokeemiliste reaktsioonide tsükkel, mis toimumiseks vajab
hapnikku. Põhifunktsioonid on orgaaniliste ühendite lõplik oksüdatsioon ja C-selgroo
andmine biosünteesireaktsioonideks.
Heksooside katabolismirajad mikroobides: glükolüüs, pentoosfosfaadirada,
Entneri-Doudoroffi rada (mis on substraadiks, mis tekib, kui palju energiat
saadakse).
– Glükolüüsi substraadid võivads olla erinevad suhkrud (glükoos, galaktoos, laktoos,
maltoos, sahharoos, fruktoos) produktiks on püruvaat. PPC rajas toimub heksooside
oksüdatsioon pentoosfosfaatide moodustumisega (Glc → riboos-P). Pentoosid on väga
olulised nukleotiidide sünteesil. Mitmetel bakteritel (näiteks Z. mobiilis, Legionella,
Pelagibacter ubiqe jpt.) puudub fosfofruktokinaas ja seetõttu ei saa töötada glükolüüs.
Neil bakteritel töötab selle asemel ED rada. substaat glükoos, produkt püruvaat ja
saagis ühe glükoosi molekuli kohta on 1 ATP, 1 NADPH ja 1 NADH.
Pentoosfosfaadiraja olulisus anabolismis.
– Annab reduktiivjõudu (NADPH) biosünteesireaktsioonideks (sh N2 fikseerimise,
sulfaadi redutseerimise ja nitraadi redutseerimise jaoks nende assimileerimisel);
Tekitab pentoose nukleiinhapete sünteesiks; Vaheproduktina moodustub rajas
erütroos-4-P, millest lähtub aromaatsete aminohapete biosüntees. Pentooside
kasutamine C-allikana toimub PPC vahendusel. Pentoosid moodustuvad looduses
näiteks hemitselluloosi hüdrolüüsil (taimede rakukestad). Sedoheptuloos-7-P on vaja
lipopolüsahhariidide sünteesil.
Kuidas toimub erinevate suhkrute sisenemine glükolüüsi?
– Maltoosi katalüüsib maltaas ja regaeerib vesi ning tekib kaks glükoosi või reageerib
fosforiga ja katalüüsib maltoos fosforülaas ja tekib glükoos ja glükoos-1-fosfaat.
samamoodi tselluloosiga, reageerib fosforiga ja katalüüsib tselluloosi fosforülaas ning
tekib glükoos-1-fosfaat ja glükoos. Sahharoos reageerib veega ja katalüüsib sukroos
ning tekib glükoos ja fruktoos. Teine võimalus on et reageerib fosforiga, katalüüsib
sahharoosi fosforülaas ja tekib glükoos-1-fosfaat ja fruktoos. Laktoos reageerib veega
ja katalüüsib beeta-galaoksidaas ning tekib galaktoos ja glükoos.
Suhkrute fosforülaasid ja nende roll energeetilises metabolismis.
– Lülitavad sukrute külge fosfori, mille eemaldamisel saadakse energiat, aga külge
panemisel kulutatakse energiat (1 ATP)
Polümeersete suhkrute kasutamine mikroobidel.
– Kasutatakse glükoosiraja jaoks.
· Käärimised: etanoolkäärimine, piimhapekäärimine,
propioonhapekäärimine, võihapekäärimine
Käärimise definitsioon ja tüübid.
– Käärimine on teatud tüüpi organismide ainevahetusprotsess, mis toimub
anaeroobses keskkonnas ühenditeni, mille edasine oksüdatsioon saab toimuda ainult
hapniku osalusel. Käärimise tüübid: etanoolkäärimine, homofermentiivne
piimhapekäärimine, heterofermetiivne piimhapekäärimine, propioonhapekäärimine,
segatüüpi käärimine, butaandioolkäärimine, võihapekäärimine, atsetoon-
butanoolkäärimine, homoatsetaatkäärimine.
Käärimis te seos heksooside katabolismi radadega.
– Homofermentatiivsel käärimisel heksoosid kataboliseeritakse glükolüüsirajas,
käärimisproduktiks on ainult piimhape ja gaase ei teki. Heterofermentatiivsel
käärimisel heksoosid kataboliseeritakse oksüdatiivses pentoosfosfaadirajas, lisaks
piimhappele moodustub ka teisi produkte (CO2). Anaeroobidel toimub püruvaadi
oksüdeerimine ferredoksiiniga (Fd). Neil puudub NAD- seoseline püruvaadi
dehüdrogenaasikompleks. Fd on mitteheemne rauda sisaldav valk. Fd- seoseline
oksüdeerimisreaktsioon esineb näiteks klostriididel käärimisradades
(võihapekäärimine). Ac-CoA-st moodustuvad käärimisproduktid, nt atsetaat.
Käärimistes moodustuvad produktid.
– etanoolkäärimine: etanool, CO2. homofermentatiivne piinhapekäärimine: laktaat.
heterofermentatiivne piimhapekäärimine: laktaat, etanool, CO2.
propioonhapekäärimine: propionaat, atsetaat, CO2. segatüüpi käärimine: etanool,
laktaat, atsetaat, suktsinaat, formaat, CO2, H2. butaandioolkäärimine: etanool,
butaandiool, laktaat, formaat, CO2, H2. võihapekäärimine: butüraat, CO2, H2.
Atsetoon-butanoolkäärimine: butanool, atsetoon, CO2, H2. homoatsetaatkäärimine:
atsetaat.
Louis Pasteur’i panus käärimiste uurimisse.
– Avastas võihappekäärimise. “käärimine on elu ilma hapnikuta.”
Kääritajate levik looduses ja nende koostöö teiste mikroobidega.
– elavad hapnikuvabades keskkondades, maapõies, meres, soolestikus.
Käärimise energiasaagis, võrdlus teiste mikroobsete katabolismiprotsesside
energiasaagisega.
– Homofermentatiivsel käärimisel 3 ATP-d ühe glükoosi molekuli kohta.
Heterofermentatiivsel käärimisel 1 ATP 1 glükoosi molekuli kohta. Propioonhappe
käärimisel tekib üks ATP. Energia saadakse substraatsest fosforüülimisest.
Biosünteesiks kasutatakse ära vaid osa energiast – rakud paljunevad aeglaselt!
Elektroni doonoriks ja aktseptoriks on orgaaniline ühend ja ATP moodustub
substraatsel fosforüülimisel.
Millised on käärimistes moodustuvad makroergilised substraadid?
– ATP, PEP, atsetüülfosfaat (Ac-P) ja atsetüül-CoA (Ac-CoA)
Milliseid aineid saab kääritada?
– suhkruid sisaldavaid aineid.
Etanoolkäärimine pärmidel.
– Pärmidel moodustub etanool glükolüüsis tekkinud püruvaadist: 1 glükoosi molekulist
moodustub 2 molekuli püruvaati, mis dekarboksüülitakse püruvaadi
dekarboksülaasiga 2 atseetaldehüüdi ja 2 CO2 molekuliks. Atseetaldehüüd (elektroni
aktseptor) redutseeritakse NADH-ga etanooliks. Redutseerimiseks kasutatav NADH
moodustub glükolüüsil. Seega on pärmide etanoolkäärimise produktideks etanool ja
süsihappegaas.
Glütserooli teke etanoolkäärimisel, selle suurendamise võimalused.
– Glütserooli teke on eriti oluline käärimise algfaasis, kui keskkonnas on palju suhkrut
(kõrge osmootne rõhk) ja kasv on kiire. Kui käärimiskeskkonnas on aineid, mis seovad
H-aktseptori atseetaldehüüdi, siis suureneb glütserooli moodustumine. Sel puhul
toimib ainsa H-aktseptorina DAP ja moodustub palju glütserooli. Etanooli ei teki või
tekib vähe. Kui keskkonda lisada neutraliseerivaid aineid (nt NaHCO3), siis toimub
Cannizarro reaktsioon – atseetaldehüüd dismuteerub atsetaadiks ja etanooliks. Sellisel
puhul toimub NADH reoksüdatsioon DAPi kaudu ja moodustub glütserool. Glükoos →
atsetaat + etanool + glütserool + CO2
Õlle tegemise põhimõte ja õllesaastajad bakterid.
– Õlle tegemise etapid: Terade idandamine (odra linnastamine), mille käigus niisutatud
terades aktiveerunud amülolüütilised ensüümid muudavad teratärklise kääritatavateks
suhkruteks (maltoosiks, maltotrioosiks, dekstriinideks, glükoosiks). Õllepärmid ise ei
suuda tärklist hüdrolüüsida! Ka proteaasid aktiveeruvad terade idandamisel ja terade
valgud hüdrolüüsitakse aminohapeteks. Idandatud terad kuivatatakse ja jahvatatakse
linnasejahuks. Virde valmistamine. Virde temperatuuri tõstetakse etapiviisiliselt. Kuna
alfa- ja glükoamülaasil on erinevad Tmax väärtused, siis sõltub sellest, kui kaua ja
millisel temperatuuril virret hoitakse see, kui palju moodustub virdesse maltoosi,
maltotrioosi, pikemaid dekstriine ja glükoosi. Sellest omakorda sõltub lõpuks valminud
õlle maitsebukett. Virde keetmine, filtreerimine ja jahutamine. Keetmisel lisatakse
humalaid. Humala parkained annavad maitset, sadestavad valke ja on
baktseritsiidsed. Enne pärmi lisamist virret aereeritakse, et sissekülvatav pärm saaks
hakata paljunema (toimub 3-4 kordne massi juurdekasv. Hapnik on vajalik ka
membraansete steroolide sünteesiks. Inokuleerimine pärmiga ja kääritamine
(hapnikku enam ei anta), mille käigus moodustub etanool ja CO2.
Käärimistemperatuur on 12°C). Käärimisel kasutatakse esmalt ära glükoos, sahharoos
(lisatakse virdele suhkrusisalduse tõstmiseks) ja maltoos. Seejärel maltotrioos ja
dekstriinid. Osa dekstriine jääb kääritamata ja need on olulised õllevahu
moodustumisel. Tänapäeval toimub kääritamine silinderkoonilistes tankides 7 päeva,
siis lastakse põhja kogunenud (flokuleerumine) pärm välja ja õlu laagerdub samas
tankis veel 3-4 päeva. Õlle filtreerimine läbi räniliivafiltrite ning pastöriseerimine 60°C.
Õlle villimine. Õlu võib tegemise käigus saastuda piimhapebakteritega (Lactobacillus
plantarum, L. brevis, Pediococcus cerevisiae), mis muudab õlle limaseks ja hapuks
(piimhape). Taluvad nii humalaid kui ja etanooli. Äadikhapebakterid (Acetobacter)
saastavad hapnikuga kokkupuutuvat õlut ja rikuvad õlle maitse (oksüdeerivad etanooli
äädikhappeks).
Õhukindlalt villitud õlut saastavad anaeroobsed bakterid Megaspaera cerevisiae ja
Pectinatus cerevisiiphilus, kes toodavad võihapet, äädikhapet, propioonhapet ja
väävelvesinikku, rikkudes õlle maitse. Kergemini saastub madala alkoholisisaldusega
õlu.
Etanooli teke bakteriaalsetes käärimistes.
– Bakteritest moodustavad etanooli käärimise kõrvalproduktina: heterofermentatiivsed
piimhapebakterid, võihapebakterid, enterobakterid (formiaatkääritajad), põhilise
käärimisproduktina moodustab etanooli Zymomonas mobilis. Etanooli moodustatakse
kas atsetüülfosfaadist, Ac-CoAst või atseetaldehüüdist redutseerimisel NADH-ga.
Milliseid radu kasutades moodustub etanool 1) pärmidel, 2) Zymomonas’el ja
3) teistel bakteritel (nt enterobakteritel ja võihapebakteritel)?
– 1) püruvaat dekarboksüülitakse püruvaadi dekarboksülaasiga 2 atseetaldehüüdi ja 2
CO2 molekuliks. Atseetaldehüüd (elektroni aktseptor) redutseeritakse NADH-ga
etanooliks.
2) püruvaat sünteesitakse atseetaldehüüdiks ja süsihappegaasiks ning see
redutseeritakse NADH2 abil etanooliks.
3) AcCoA redutseeritakse NADH2 juuresolekul atseetaldehüüdiks ja sealt samamoodi
etanooliks.
Mis on Zymomonas mobilis’e eelis etanoolkääritajana?
– Zymomonas kasvab anaerobioosis palju kiiremini, kui Saccharomyces. Peaaegu kogu
substraadi konverteerib etanooliks, rakumassi moodustub vähe. Membraani
stabiliseerimiseks ei vaja hapniku juuresolekut – hopanoidide süntees ei vaja
hapnikku. Talub väga kõrget suhkrusisaldust söötmes (kuni 30-40%) ja etanooli
taluvus on tal ka kuni 13%, mis on bakterimaailmas haruldane. Enamus baktereid
talub vaid 1-2% etanooli.
Millised mikroobid ja toorained sobivad bioetanooli tootmiseks?
– Bioetanooli tootmisel kasutatakse kääritajana peamiselt Saccharomyces’t. Kuid
taimsed jäätmed sisaldavad lisaks tselluloosile ka hemitselluloosi, mille hüdrolüüsil
vabanevad pentoosid, mida Saccharomyces kääritada ei suuda. Kääritatavad jäätmed
võiks olla: suhkruroo jäätmed ehk bagass; õled; saepuru jmt. Toodetakse etanooli ka
suhkruroost (sahharoos) Zymomomas mobilis’e abil.
Millised on piimhapekäärimise tüübid?
– Homo- ja heterofermentatiivne.
Homo- ja heterofermentatiivsed piimhapebakterid: mis on erinevused ja
sarnasused.
– Suhkrud lagundatakse glükolüüsirajas püruvaadini. Püruvaat redutseeritakse laktaadi
dehüdrogenaasiga laktaadiks. Sellega reoksüdeeritakse glükolüüsis moodustunud
NADH. Gaase ei teki. ATP saagis 2 mooli 1 mooli Glc kohta. Obligaatselt
heterofermentatiivsetel piimhapebakteritel on defektne glükolüüs. Nt puudub
fruktoosdifosfaadi aldolaas. Seetõttu lagundavad nad heksoose PPC abil.
Heterofermentatiivsel käärimisel moodustub lisaks piimhappele veel etanooli,
atsetaati ja CO2. Summaarne ATP saagis on heterofermentatiivsel käärimisel poole
väiksem, kui homofermentatiivsel (1 ATP 1 molekuli kääritatud glükoosi kohta).
Seetõttu toodavad heterofermentatiivsed piimhapebakterid sama koguse suhkru
kääritamisel poole vähem rakumassi kui homofermentatiivsed.
Bifidobakterid ja vastav käärimine.
– Bifidobakteritel on omapärane käärimine. Käärimine meenutab heterofermentatiivset
piimhapekäärimist, sest osalevad fosfoketolaasid. Bifidobacterium kääritab kaks mooli
glükoosi 2 mooliks piimhappeks ja 3 mooliks äädikhappeks. Protsessi võtmeensüümiks
on Fru- 6-P fosfoketolaas. Fruktaanide (inuliini) hüdrolüüsimise jaoks on
eksoinulinaasid, mis tekitavad inuliinist fruktoosi, mida kääritada.
Kuidas valmivad hapupiimatooted ja millised piimhapebakterid seal
esinevad?
– Piinhapebakterid on laktokokid, streptokokid, laktobatsillid, bifidobakterid,
pediokokid, oenokokid. Valmivad piimatoodete kääritamisel piimhapebekteritega.
Propioonhapekäärimine, selle produktid ja mõned näited vastavatest
bakteritest.
– Propioonhappebaktereid on palju mäletsejate loomade (lehm, lammas) vatsas.
Vatsas toimub taimsete polüsahhariidide kääritamine rasvhapeteks bakterite
vahendusel. Propioonhappebakterid kääritavad vatsas edasi ka piimhapebakterite
poolt moodustatud piimhapet propionaadiks. Propionaat on põhiline
glükoneogeneetiline substraat mäletsejal loomal (sellest sünteesitakse maksas
piimasuhkur ehk laktoos). Inimestel on propionaat ka glükonegeneetiline. Produktideks
on propionaat, atsetaat, CO2. Propionibacterium shermanii, Cutibacterium acnes,
Clostridium propionicum ja Bacteroides.
Membraanne fosforüülimine propioonhapekäärimise rajas.
– Püruvaadist karboksüülimise ja edasiste muundumiste kaudu moodustub propionaat.
Protsessis osaleb ka membraaniga seotud fumaraadi reduktaas. Kui fumaraat
atsepteerib elektronid NADH-lt, siis kaasneb sellega prootonite liikumine läbi
membraani ja membraanne ATP süntees (membraanne fosforüülimine). See tõstab
propioonhapekäärimise energeetilist efektiivsust.
Võihapekäärimine, selle produktid.
– Võihapet moodustavad käärimisel reeglina ainult ranged anaeroobid. Perekonnad
Clostridium (C. butyricum, C. acetobutylicum), Butyrivibrio, Eubacterium,
Fusobacterium, Lachnospira, Roseburia, Faecalibacterium. Käärimisel moodustub
gaase, atsetaati ja orgaanilisi lahusteid: etanooli, butanooli, atsetooni, isopropanooli.
Jämesooles stimuleerib butüraadi moodustumist ka käärimistes moodustuv atsetaat –
see konverteeritakse teiste bakterite vahendusel butüraadiks. Klostriidid lagundavad
heksoose glükolüüsirajas.
Neutraalsete produktide (atsetoon, butanool jne) teke.
– Tekivad atsetoon-butanoolkäärimise, võihappekäärimisel ja propioonhapehäärimisel.
Milliseid käärimisi viivad läbi klostriidid?
– Etanoolkäärimist, võihappekäärimist,
Aminohapete paariskääritamine klostriididel (Sticklandi reaktsioon).
– 1934. a. avastas Stickland, et Clostridium sporogenes ei suuda Ala ja Gly eraldi
kääritada, kuid koos (Ala+ Gly) suudab. Ala toimib H-doonorina ja Gly aktseptorina.
Üks aminohape oksüdeeritakse ja teine redutseeritakse. ATP saadakse substraatsel
fosforüülimisel.
Bakteritevaheline vesiniku ülekanne ja selle tähtsus mikroobidele.
– Üks bakter eritab vesinikku ja teine tarvitab seda. Protsess on looduses väga levinud,
sest see on kasulik mõlemale partnerile. Bakteritevaheline H2 ülekanne võimaldab
kääritajal bakteril rohkem ATP-d sünteesida. Käärimistes moodustunud NADH-d
võimalik reoksüdeerida Fd-ga ja redutseeritud ferredoksiini omakorda oksüdeerida
gaasilise vesiniku eraldumisega. See on bakteritele energeetiliselt kasulik. Bakter saab
toota rohkem ATP-d. Protsess saab toimuda vaid siis, kui väliskeskkonnas hoitakse H2
kontsentratsioon väga madalal. Keskkonnas peab olema mikroobe, kes aktiivselt
vesinikku tarbivad (metanogeene, anaeroobseid hingajaid).
Formiaatkäärimine ja selle produktid.
– Bakterid, kes moodustavad käärimisel formiaati (HCOOH; sipelghape). Kuna aga
formiaati teistes käärimisradades ei moodustu, nimetatakse sellist hapete segu
tekitavat käärimist formiaatkäärimiseks (segahape käärimine). Formiaatkäärimine on
iseloomulik enterobakteritele.
Püruvaadi võimalikud katabolismi teed enterobakteritel.
– redutseeruda piimhappe tekkega, laguneda Pyr:formiaatlüaasi reaktsioonis
formiaadiks ja AcCoA-ks, PEP võib karboksüüluda oksaalatsetaadiks ja sellest
moodustub suktsinaat. Osaleb fumaraatses hingamises (membraanne fosforüülimine).
Käärimisproduktide moodustumine püruvaadist.
– Kaks tüüpi: hapete segu teke: iseloomulik perekondadele Escherichia-Salmonella-
Shigella, iseloomulik püruvaadist atsetaadi jt hapete teke, butaandioolkäärimine:
iseloomulik perekondadele Erwinia-Enterobacter-Serratia, püruvaadist moodustatakse
neutraalset butaandiooli.
· Anaeroobne hingamine
Anaeroobne hingamine, selle definitsioon ja levik looduses.
– Anaeroobne hingamine – protsess, kus oksüdeeritavalt substraadilt (orgaanilised või
anorgaanilised ühendid) ärastatud elektronid kantakse hapnikust erinevale valdavalt
anorgaanilisele elektroni aktseptorile ja ATP moodustub membraansel fosforüülimisel.
Anaeroobid on kõik peale aeroobsete bakterite, seente, algloomade ja eukarüootide.
Anaeroobse hingamise tüübid: mis on iga tüübi puhul elektroni
lõppaktseptor ja moodustunud redutseeritud produkt.
– prootonite redutseerimine - H+->H2. Atsetogenees - CO2->CH3COO-.
Väävlihingamine - S->HS-. Metanogenees - CO2->CH4. Sulfaatne hingamine ->
SO3(2-)->HS-. Fumaraatne hingamine - fumaraat->suktsinaat. Rauahingamine - Fe3+-
>Fe2+. Nitraatne hingamine - NO3- -> NO2-. Denitrifikatsioon O2->H2O
Kuidas moodustub ATP anaeroobsel hingamisel ja palju seda tekib?
– 1 ATP ae4roobse hingamise 3 ATP vastu.
Kus anaeroobsed hingajad looduses elavad?
– Mere ja maapinna sees, kus pole hapnikku lähedal. Inimese soolestikus.
Milliste mikroobidega elavad koos anaeroobsed hingajad ja miks?
– Elavad koos kääritajatega, sest nad oksüdeerivad edasi kääritavate mikroobide
toodetud substraate – orgaanilisi happeid ja alkohole ning vesinikku.
Nitraatne hingamine ja denitrifikatsioon.
– Nitraatne hingamine on protsess, kus oksüdeeritavatelt (orgaanilistelt) ühenditelt
ärastatud elektronid kantakse nitraadile, mis redutseerub nitritini ja eritatakse
keskkonda. ETA lõpplüli on nitraadi reduktaas.
Nitraadi ammonifikatsioon.
– Nitraadiga hingajatel võidakse moodustunud nitrit edasi ammoniaagini redutseerida
energiat mitte andvate reaktsioonide vahendusel. Nitraat → Nitrit → HNO → NH2OH →
NH3. Bakteril on sellest kasu, sest nitrit on toksiline (mutageen) ja bakterid saavad
nitritit redutseerides reoksüdeerida orgaaniliste ainete lagundamisel moodustunud
NADH2- d. Sel juhul on neil võimalik toota enam ATP-d metabolismis moodustunud Ac-
CoA-st (substraatne fosforüülimine).
Milles seisneb nitraatide toksilisus imikutele ja täiskasvanutele?
– Imikutel, kellel pole välja arenud mikroobikooslus välja arenenud, imenduvad nitritid
verre ja seostuvad hemoglobiiniga ja oksüdeerivad raua hemoglobiinis, moodustub
methemoglobiin, mis ei suuda hapnikku transportida. Ohustatud on ka alahappelise
maoga täiskasvanud.
Sulfaatne hingamine. Sulfaatsete hingajate eelistatud oksüdeeritavad ained.
– Sulfaatsel hingamisel (dissimilatoorne redutseerimine) aga kasutatakse sulfaati
ainete oksüdeerimisel terminaalse elektroniaktseptorina ja moodustuv H2S eritatakse
rakust välja. Eelistatud ained on SO4(2-), sulfaat. Anaeroobsete hingajate
meelissubstraadid on atsetaat, laktaat, püruvaat, malaat ja etanool. Need
moodustuvad käärimistes. Sulfaatsel hingamisel võidakse lagundada ka alkaane ja
aromaatseid ained (fenool, bensoaat, toluool).
Väävlihingamine. Fumaraatne hingamine. Rauahingamine.
– Väävelhingamine: elektronide lõppaktseptor hingamisel on väävel. Produktina tekib
väävelvesinik. Väävlihingajad on näiteks Desulfuromonas acetoxidans ja Pyrodictium
occultum, Desulfurococcus, Pyrococcus, Thermococcus jt. Fumaraatsel hingamisel
kantakse ainete oksüdatsioonil ärastatud elektronid fumaraadile, mis redutseeritakse
suktsinaadiks. Kompleksis osaleb fumaraadi reduktaas. Sel teel moodustub suktsinaati
paljudel anaeroobsetel bakteritel loomavatsas (nt Wolinella) ja ka jämesooles (nt
Bacteroides). Rauahingamine on levinud mullabakteritel. Paljud rauahingajad on
võimelised hingama ka mangaaniga. Kõige rohkem on rauahingamist uuritud bakteril
Shewanella. Shewanella oneidensis saab hingata hapnikuga, nitraadiga, DMSO-ga, aga
ka väga paljude metallidega (rauaga, mangaaniga, uraaniga jne).
Kuidas toimub rakuväline elektronide ülekanne rauabakteritel nanojuhtmete
abil?
– Kuna Fe(III) ja Mn(IV) oksiidid on lahustumatud, siis saab raua ja mangaani
kasutamine (redutseerumine) toimuda vaid väljaspool rakku. Elektronid suunatakse
rakust välja rauale nanojuhtmeid mööda. Raud toimib nagu anood. Nanojuhtmed
saavad seostuda ka elektroodiga (anoodiga) ja anda elektronid sinna. Tekib vool. Raud
ja mangaan muutuvad redutseerumisel lahustuvaks ja liikuvaks.
Prootonite redutseerimine arhel Pyrococcus furiosus kui ürgne anaeroobse
hingamise viis.
– Orgaanilised ained oksüdeeritakse Fd-seoseliselt. Membraanne hüdrogenaas
oksüdeerib
redutseeritud Fd prootonitega. Moodustub gaasiline H, osa prootoneid suunatakse läbi
membraani prootongradiendi tekkega. ATP süntaas sünteesib prootongradiendi abil
ATP-d. Glükolüüsis toimuvad Fd-seoselised oksüdatsioonireaktsioonid (ürgne
glükolüüs).
Milles seisneb metanogeneesi eripära?
– Elektroni doonorina (oksüdeeritava substraadina) sobivad metanogeenidele vesinik,
formiaat, atsetaat jmt ühendid ning elektroni lõppaktseptoriks on CO2, mis
redutseeritakse metaanini. Protsessi käigus toimub membraanpotentsiaali
genereerimine ja selle arvel ATP süntees.
· Kemolitotroofia
Kemolitotroofia mõiste ja avastaja (Sergei Vinogradskiy)?
– Kemolitotroofia on toitumistüüp, kus mikroobid saavad energiat anorgaaniliste
ühendite oksüdatsioonist. Anorgaanilistelt ühenditelt ärastatud elektronid kantakse
ETA vahendusel enamasti hapnikule ning membraanil moodustuva prootongradiendi
arvel sünteesitakse membraansel fosforüülimisel ATP.
Kuidas toimub ATP süntees kemolitotroofidel?
– Anorgaanilistelt ühenditelt ärastatud elektronid kantakse ETA vahendusel enamasti
hapnikule ning membraanil moodustuva prootongradiendi arvel sünteesitakse
membraansel fosforüülimisel ATP.
Kemolitotroofsed bakterite põhirühmad.
– Nitrifitseerijad (oksüdeerivad ammoniaaki või nitritit), tioonbakterid (väävlibakterid)
(oksüdeerivad redutseeritud S-ühendeid), rauabakterid (oksüdeerivad Fe(II)),
vesinikubakterid (oksüdeerivad vesinikku), vingugaasibakterid (oksüdeerivad CO-d).
Nitrifikatsioon, selle kaks etappi. Millistel tingimustel on nitrifikatsioon
inhibeeritud?
– Nitrifikatsioon – ammoniaagi oksüdatsioon nitraadiks on kahe-etapiline. Kumbagi
etappi
viivad läbi erinevad bakterid. Esimene etapp esineb ka arhedel! I etapp: NH3 -> NO2-
II etapp: NO2- ->NO3-. INhibeeritud happelises keskkonnas ja hapnikuvabas
keskkonnas.
Miks, kuidas ja millal toimub nitrifikatsioon ja denitrifikatsioon
veepuhastuses?
– Org. aine ammonifikatsioonil tekib palju NH3. Nitrifitseerijaid on palju reovees. Kui
ammoniaagirikas reovesi (kalatiikide vesi, olmereovesi) juhtida looduslikesse
veekogudesse ilma eelneva puhastuseta, siis kulub veekogus ammoniaagi
oksüdeerimiseks palju hapnikku (suur bioloogiline hapnikutarve). Seega tuleks reovesi
puhastada (ammoniaak ära oksüdeerida) enne loodusesse juhtimist. Heitvee
aeroobsel puhastusel oksüdeerub ammoniaak nitrifitseerijate bakterite vahendusel
nitritiks ja nitraadiks. Nitraat ja nitrit redutseeritakse edasises heitvee anaeroobse
töötlemise käigus gaasilisteks produktideks. Osalevad denitrifitseerijad bakterid.
Seega on veepuhastamisel lämmastikuühenditest mitu etappi, esimesed etapid on
aeroobsed, viimane anaeroobne.
Kuidas toimub redutseeritud väävliühendite oksüdatsioon ja millised on
tioonbakterid?
– Sergei Vinogradski kirjeldas aastal 1887 niitja tioonbakteri Beggiatoa, kes saab
energiat H2S oksüdatsioonist, mille tulemusena kogunevad rakkudesse ajutiselt
väävliterad. Tegi kindlaks, et see bakter saab süsinikuallikana kasutada CO2. Seega
tegi Vinogradskiy esimesena kindlaks kemolitoautotroofse toitumistüübi bakteritel.
Väävli oksüdeerijad peavad selleks kinnituma väävliosakeste pinnale. Sulfiidilt antakse
elektronid flaviinile, teistelt S-ühenditelt aga tsütokroomidele. Elektronide
lõppaktseptor on hapnik. Väävliühendite oksüdatsioonil moodustub väävelhape.
Mille poolest on tuntud Thiomargarita ja Thioploca?
– Mõlemad on tioonbakterid: Thiomargarita on ülisuur bakter nitraadivakuoolidega,
Thioplocal on nitraadivakuoolid.
Mis roll on tioonbakteritel toru-usside toitekoes?
– Hüdrotermaalsete lõõride asukate toru-usside (tube worms) kehas on spetsiaalne
kude (trofosoom), kus elavad H2S oksüdeerivad autotroofsed bakterid, kes varustavad
ussi orgaanilise ainega. Väliskeskkonnast lõpuste kaudu jõuab ussi kehasse hapnik,
CO2 ja H2S, millest trofosoomis elavad bakterid sünteesivad orgaanilist ainet.
Vesiniku ja vingugaasi oksüdatsioon, vastavad bakterid.
– Vesinikubakterid kuuluvad perekondadesse Ralstonia, Paracoccus, Aquaspirillum,
Xanthobacter, Rhizobium, Nocardia, Mycobacterium, Bacillus. Bakterid Aquifex ja
Hydrogenobacter on vesinikubakterid. Lisaks vesinikule saavad nad energiaallikana
kasutada ka väävlit ja tiosulfaati. Saavad energia H2 oksüdatsioonist ja C-allikana
kasutavad CO2. Nad on kõige kiirema kasvuga kemolitoautotroofid. Termofiilsetel
liikidel võib generatsiooniaeg olla 1 tund. 6H2 + 2O2 + CO2 -> (CH2O) + 5H2O.
Vingugaasibakterid on Pseudomonas carboxydovorans, Seliberia, Comamonas,
Achromobacter, Arthrobacter, Azomonas, Azotobacter, Alcaligenes, Streptomyces.
eroobsed bakterid, kes saavad kasvada CO kui ainsa C- ja energiaallika arvel. 2 CO +
O2 -> 2 CO2.Ainult 4% oksüdeeritavast CO-st lülitub rakuainesse ja 96% sellest läheb
energeetiliseks otstarbeks.
Raua oksüdatsioon, vastavad bakterid.
– Raud vabaneb huumusest selle lagunemisel, oksüdeerub keemiliselt ja sadeneb
mikroobide pinnale (raku pind on negatiivselt laetud). Neutraalses keskkonnas ja
aeroobsetes tingimustes oksüdeerub Fe kiiresti keemiliselt ja sadeneb välja.
Happelises keskkonnas Fe keemiliselt ei oksüdeeru! Fe ja Mn ühendid võivad
oksüdeeruda ka mikroobide ainevahetusproduktide (H2O2) toimel. H2O2 eritatakse
rakkude kapslitesse ja neutraalses ning aluselises keskkonnas saab toimuda Fe ja Mn
keemiline oksüdatsioon H2O2-ga. Anaeroobsete fototroofsete bakterite vahendusel
(banded iron). Raua ja mangaani oksüdatsioon kemolitoautotroofsete bakterite
vahendusel. Rauabakterite hulka kitsamas mõttes (kemolitoautotroofsed rauabakterid)
kuluvad: Thiobacillus (Acidithiobacillus) ferrooxidans, Leptospirillum ferrooxidans,
Sulfolobus acidocaldarius, Gallionella ferruginea, Marinibacter profundus.
Mille poolest on tuntud rauabakter Gallionella?
– G. ferruginea saab energiat nii väävliühendite kui ka raua oksüdatsioonist. Oakujulise
raku nõgusalt küljelt eritatakse välja lintjas limsaba, mis hiljem keerdub. Sabasse
ladestuvad raua oksüdatsiooniproduktid. Lima süntees ja eritamine sabana on vajalik
selleks, et rakupind ei ummistuks lahustumatute rauaühenditega. Looduses esineb
Gallionella mageveekogudes, aga ka veefiltrites aeroobse ja anaeroobse tsooni piiril.
Seal leidub nii redutseeritud S- kui ka rauaühendeid.
· Õhulämmastiku sidumine
Kuidas toimub N2 fikseerimine mikroorganismidel ja millised on tuntumad N2
fikseerivad mikroobid?
– N2 fikseerijad: Aeroobid ja anaeroobid, Bakterid ja arhed, Fotosünteesijad
(tsüanobakterid, anoksügeensed fotosünteesijad), Kääritajad, Anaeroobsed hingajad,
Kemolitotroofid, Metanogeensed arhed, Taimedega sümbioosis N2 sidujad.
Mikroorganismides kulub N2 redutseerimiseks NH3-ni palju energiat (16-24 ATP
molekuli ühe molekuli redutseeritava N2 molekuli kohta). Energia saadakse keemiliste
ühendite oksüdatsioonist või valgusenergia arvel. Nitrogenaaskompleks koosneb
kahest valgust: dimeerne nitrogenaasi reduktaas, mis sisaldab rauda, ja tertrameerne
nitrogenaas, mis sisaldab nii rauda kui ka molübdeeni. Nitrogenaasi reduktaas
redutseeritakse ferredoksiiniga. Nitrogenaasi reduktaas redutseerib seejärel
nitrogenaasi, mis kannab elektronid ja prootonid õhulämmastikule (N2).
Erinevad võimalused, kuidas nitrogenaaskompleksi hapniku eest kaitsta.
– Azotobacter’il (aeroobne mullabakter) on väga aktiivne hapnikuhingamine, mis hoiab
rakus hapniku sisalduse madalal. N2 fikseerimine toodab kõrvalproduktina vesinikku.
Selle oksüdeerimine hapnikuga hüdrogenaasireaktsioonis kaitseb nitrogenaasi.
Limakapsel takistab hapniku difusiooni rakuni. Näiteks kui Azotobacter’it kasvatada
õhu käes, kus on “tavaline” hapnikusisaldus (21 %), sünteesib ta endale paksu
limakapsli, kui aga hapnikku on vähe (2.5 %), siis kapslit ei sünteesita. Niitjatel
tsüanobakteritel on heterotsüstid, milles nitrogenaas on hapniku eest kaitstud.
Fakultatiivsed anaeroobid fikseerivad N2 ainult anaeroobsetes tingimustes.
Rhizobium’il (sümbiontsetel mügarbakteritel) kontrollib hapniku difusiooni bakteroidini
leghemoglobiin. Looduses kaitsevad aeroobseid N2 fikseerijaid liigse hapniku eest
teised bakterid, kes hapnikku aktiivselt kasutavad.
Mis on leghemoglobiin?
– hemoglobiini sarnane pigment, mis on vajalik hapnikusisalduse kontrolliks
mügarakoes. Ta seob hapnikku ja vabastab seda väga aeglaselt. Leghemoglobiiniga
seotud ja vaba hapniku hulkade suhe juuremügaras on 10 000:1.
Hüdrogenaas ja selle roll.
– kiiresti oksüdeerib moodustuva vesiniku hapnikuga veeks. Sellega hoitakse
nitrogenaasi ümbrus hapnikuvaene.
Vabaltelavad ja sümbiontsed N2 fikseerijad.
– Anaeroobsetest N2 fikseerijatest bakteritest kirjeldati esmalt Clostridium
pasteurianum. Suhkrurootaime kudedes on palju Acetobacter diazotrophicus’t, kes
fikseerib seal N2, kasutades energiaallikana taimemahla sahharoosi. Veekogudes on
väga olulisteks N2 fikseerijateks tsüanobakterid Nostoc, Anabaena, Oscillatoria jt. Ka
riisipõldudel fikseerivad tsüanobakterid N2. Tsüanobakteritel on kirjeldatud ligi sada
N2 fikseerivat liiki. Paljudel niitjatel tsüanobakteritel on niidis spetsiaalsed rakud
(heterotsüstid), kus “töötab” nitrogenaas ja on seal kaitstud hapniku eest.
Liblikõieliste taimede juuremügarates fikseerivad lämmastikku mügarbakterid.
Liblikõielistega on sümbioosis perekonna Rhizobium ja Bradyrhizobium, Azorhizobium
ja Sinorhizobium jt esindajad. Need on kõik uued perekonnad, mis on moodustatud
vanast Rhizobium perekonnast. Kuuluvad peamiselt alfa-proteobakterite hulka.
Rhizobium fikseerib N2 bakteroidina. Bakteroidid on erilised diferentseerunud
bakterirakud, mis moodustuvad juuremügaras vegetatiivsetest rakkudest.
Mis on heterotsüstid tsüanobakteritel ja millised protsessid neis toimuvad?
– Tsüanobakterid on aeroobsed fotosünteesijad bakterid, kellel fotosünteesi
tulemusena moodustub hapnik. Seega peavad neil olema võimalusi, kuidas
nitrogenaasi kaitsta rakus moodustuva hapniku eest. Paljudel niitjatel tsüanobakteritel
on niidis spetsiaalsed rakud (heterotsüstid), kus “töötab” nitrogenaas ja on seal
kaitstud hapniku eest. Heterotsüst ja akineet. Heterotsüstid moodustuvad
tsüanobakteri niiti siis, kui keskkonnas on vähe seotud lämmastikku (nitraat- ja
ammooniumlämmastikku). Siis diferentseerub 5-10% niidirakkudest heterotsüstideks.
Heterotsüstid ei tooda hapnikku ega fikseeri CO2! Neis ei ole karboksüsoome.
Heterotsüst ei tooda hapnikku, kuna seal puudub fotosüsteem II ja ei toimu vee
fotolüüsi. N2 difundeerub heterotsüsti naaberrakkudest. N2 redutseerimisel
moodustunud ammoonium lülitatakse glutamiini koosseisu ja transporditakse
heterotsüsti naaberrakkudesse. Heterotsüstis on anaeroobne keskkond ja seal toimub
N2 redutseerimine. Kasutatakse valgusenergiat.
Mügarbakterid ja juuremügara teke.
– Taimejuured eritavad flavonoide, mis meelitavad kohale mügarbakteri. Mügarbakter
eritab vastuseks nod-faktoreid (kitiinilaadsed oligosahhariidsed ühendid ehk
lipokitooligosahhariidid), mille järel juurekarv kõverdub kasvades ja bakter hakkab
tungima taimekoesse. Infektsiooniniit on tselluloosiga vooderdatud torutaoline kanal
juurekoes, mille kaudu toimub taimekudede nakatumine mügarbakteritega. Bakterid
jagunevad selles kanalis ja liiguvad edasi taimejuure kudedesse. Bakteri
signaalmolekulide toimel hakkavad juurekoe rakud jagunema, neisse tungivad
bakterid ja moodustub juuremügar.
· Bakteriaalne fotosüntees
Bakteriaalne fotosüntees, selle eripära.
– Valguse toimel lüüakse fotosünteesipigmendist välja elektron
(oksüdatsioonireaktsioon). Elektronid liiguvad membraanses ETA-s ja sellega kaasneb
membraanne fosforüülimine. ATP-d ja reduktiivjõudu kasutatakse CO2
redutseerimiseks orgaanilisse ainesse.
Fotosünteesipigmendid ja membraansed elektrontransportahelad.
– Taimedele omane Chl a neelab sinist ja punast valgust ja peegeldab tagasi rohelist ja
kollast! Taimed on rohelised. Täiendavateks pigmentideks fotosünteesivatel bakteritel
on karotenoidid. Need on vees mittelahustuvad ühendid, värvilt on nad kollased,
punased, pruunid või rohelised. Valgust koguvad pigmendid (karotinoidid ja klorofüllid)
paiknevad membraanis. Nad suunavad valguse edasi reaktsioonitsentri klorofülli
molekulidele. Fotosünt. pigmendid paiknevad purpurbakteritel rakumembraani
torujatel sopististel. Neid on nimetatud ka kromatofoorideks.
Rohelistel bakteritel paikneb osa pigmente (valgust koguvad tsentrid) klorosoomides.
Pigmentide sisaldus rakkudes on suurim nõrga valgustatuse tingimustes ja
anaerobioosis. Valgustpüüdvat pinda saab suurendada ka jätketega. Valgusenergia
mõjul viiakse reaktsioonitsentri klorofüllist välja elektron, mis liigub
elektrontransportahelas kandjalt kandjale. Protsessiga kaasneb membraanil
prootongradiendi (proton flow) teke. Kuna kinoonid kannavad üle nii prootonit kui ka
elektroni, siis tõmmatakse elektroni seostumisega kinoonile tsütoplasmast juurde
prooton, mis hiljem suunatakse läbi membraani välja. Moodustub prootongradient
Millised on erinevad välised redutseerijad fotosünteesil?
– Purpursed mitteväävlibakterid: H2, H2S, tiosulfaat, orgaanilised ühendid, Fe2+.
Purpursed väävlibakterid: H2S, S, tiosulfaat, nitrit, Fe2+, arseen (III). Rohelised
väävlibakterid: H2S, orgaanilised ühendid. Rohelised väävlibakterid: H2S, S.
Tsüanobakterid: H2O.
Oksügeenne fotosüntees tsüanobakteritel ja anoksügeenne fotosüntees
rohelistel ja purpurbakteritel.
– Tsüanobakteritel on nii FSI kui ka FSII, mis töötavad koos. See protsess võimaldav
vee fotolüüsi. Protsessis eraldub kõrvalproduktina hapnik, mis pärineb veest!
Valgusega klorofüllist väljalöödud elektronid liiguvad primaarsele elektroniaktseptorile
FSII ETA-s. Protsessi käigus lõhustab FSII vee (2H2O → O2 + 4H+ + 4 elektroni). Need
elektronid asendavad klorofüllist väljalöödud elektrone. Elektronide liikumisega
membraanis luuakse sellele prootongradient ja sünteesitakse membraansel
fosforüülimisel ATP. Valguse toimel lüüakse FSI klorofüllist välja elektronid, mida
kasutatakse reduktiivjõu NADPH sünteesiks. See osaleb Calvini tsüklis CO2
redutseerimisel suhkruteks. Enamik tsüanobaktereid suudab fikseerida N2. Niitjatel
tsüanobakteritel toimub see heterotsüstides, kus nitrogenaaskompleks on kaitstud
fotosünteesil eralduva hapniku eest. Purpursed väävlibakterid kasutavad eksogeense
redutseerijana redutseeritud väävliühendeid. Väävliühendite oksüdatsioonil
moodustub väävelvesinikust elementaarväävel, mis on vaheproduktiks ja
oksüdeeritakse edasi sulfaadiks. Purpurse mitteväävlibakterid eelistavad orgaanilisi
ühendeid, ui kasutavad H2S fotosünteesil, siis väävliteri rakkudesse ei kogu, ei talu
kõrget H2S sisaldust, tsitraaditsükkel täielik, põhiline toitumistüüp
fotoorganoheterotroofia: orgaanilistest ühenditest kasutavad orgaanilisi happeid,
alkohole ja suhkruid. Rohelised väävlibakterid kasutavad elektronidoonorina
(redutseerijana) kasutavad väävelvesinikku, tiosulfaati, vesinikku, elementaarväävlit ja
rauda. Orgaanilisi ühendeid kasutavad ainult täiendava C-allikana. Enamik
rohebaktereid fikseerib õhulämmastikku. Rohelised mitteväävlibakterid sarnanevad
purpursete mitteväävlibakteritega orgaaniliste ühendite eelistamise poolest
(fotoorganotroofia), tsüanobakteritele niitja ehituse ja libiseva liikumise poolest ja
rohelistele S-bakteritele klorosoomide esinemise poolest.
Fototroofsete bakterite rühmad: bakterid ja nende elupaigad.
– Oksügeensed ehk hapnikku tekitavad fotosünteesijad (taimed, vetikad,
tsüanobakterid).
Kasutavad fotosünteesil CO2 redutseerijana vett, vabaneb hapnik. Anoksügeensed ehk
hapnikku mittetekitavad fotosünteesijad (kõik ülejäänud fotosünteesivad bakterid, nt
purpur- ja rohebakterid). Kasutavad redutseerijana veest erinevaid aineid, näiteks
redutseeritud S-ühendeid.
Mille poolest on tuntud Chlorochromatium aggregatum?
– koosneb kesksest heterotroofsest bakterist (beeta-proteobakter) ja teda
ümbritsevatest rohelistest väävlibakteritest.
Mis on Vinogradskiy sammas (kolonn)?
– modelleerib looduses mikroobide kihistumist ja hapniku- ja H2S gradienti veekogus.
Pinnal elavad hapnikku tootvad tsüanobakterid, nende all purpursed
mitteväävlibakterid, sügavamal anaeroobsed fototroofid – purpursed väävlibakterid ja
rohelised väävlibakterid.
Halobakterid (ehk haloarhed) kui bakterirodopsiiniga fotosünteesivad
prokarüoodid.
– Fotosünteesipigmendiks on neil bakterirodopsiin. See on pigment, kus valguga on
seotud karotinoidne ühend – retinaal. Halobakterid on fakultatiivsed fototroofid. Nad
saavad kasutada orgaanilisi aineid (nt orgaanilisi happeid) – oksüdeerivad neid
hapniku või nitraadiga. Hapniku sisalduse langedes keskkonnas lülitub rakkudes sisse
bakterirodopsiini süntees. Bakterirodopsiin toimib prootonpumbana, mis võimaldab
sünteesida ATP-d valgusenergia arvel. Bakterirodopsiin on lihtne prootonpump, tema
tööks ei ole vaja tsütokroome ega muid täiendavaid pigmente. Seega ei inhibeeri seda
pumpa ka tsüaniid ega asiid.
Kuidas töötab bakterirodopsiin lihtsa prootonpumbana?
– Hapniku sisalduse langedes keskkonnas lülitub rakkudes sisse bakterirodopsiini
süntees. Bakterirodopsiin toimib prootonpumbana, mis võimaldab sünteesida ATP-d
valgusenergia arvel. Bakterirodopsiin on lihtne prootonpump, tema tööks ei ole vaja
tsütokroome ega muid täiendavaid pigmente. Seega ei inhibeeri seda pumpa ka
tsüaniid ega asiid.
fotoorganoautotroofe ei esine looduses
Kemolitotroofia esineb prokarüüotidele ainuomane
Metanogenees omane ainult arhedele
Moodle testid:
Mikrobioota ja antibiootikumid
1. Märgi millised antibakteriaalseid komponente inimene ise sünteesib
– Lüsosüüm ja happed
2. Nimeta kaks bakteriperekonda, kelle hulgas on antibiootikumide sünteesijaid
– Streptomyces, Bacillus
3. Ühenda omavahel mõiste ja sünonüümid
– kõik mikroobid, nende aktiivsused ja komponendid - mikrobioom
mikroobne metagenoom - mikrobioom
mikrofloora - mikrobioota
mikroobikooslus - mikrobioota
4. Märgi õiged väited soolestiku mikrobioota ja selle mõjude kohta
– Helicobacter pylori saab maos ellu jääda ja paljuneda tänu uraaside toimele
Probiootikumid on tervisele kasuliku mõjuga bakterid
5. Märgi tunnused, mille poolest erinevad gnotobiondid tavatingimustes kasvatatud
loomadest
– Õhem sooleseid, vähem metaboliite veres, nõrgem immuunsüsteem
6. Ühenda omavahel antibiootikumide grupp ja selle märklaud bakterirakus
– Vankomütsiin - rakukest
Tsiprofloksatsiin - nukleiinhapete süntees
penitsiliinid - rakukest
rifampiin - nukleiinhapete süntees
makroliidid - valgusüntees
trimetoprim - foolhappe süntees
klooramfenikool - valgusüntees
karbapeneemid - rakukest
streptomütsiidid - valgusüntees
sulfaniilamiidid - foolhappe süntees
tetratsükliin - valgusüntees
7. Reasta bakterid alates WHO andmetel kõige ohtlikumast patogeenist
– Tuberkuloositekitaja Mycobacterium tuberculosis
Acinetobacter baumanii
Staphylococcus aureus (MRSA)
Haemophilus influenzae
Treponema pallidum
8. Märgi õiged väited inimese normaalse mikrobioota kohta
– Erinevatel inimestel on samades kehapiirkondades sarnased mikroobid
Mikroobide mitmekesisuse vähenemine võib põhjustada haiguslikke protsesse
Normaalse mikrobioota koosseis muutub inimese elu jooksul palju
Veri, ülemised kusetees ja alumised hingamisteed on organismis tavatingimustes
mikroobivabad
9. Millised on levinud antibiootikumide resistentsusmehhanismid bakteritel? Märgi kõik
sobivad
– Kõik mainitud (märklaua muutmine, antibiootikumi rakust välja pumpamine,
märklaua ülehulgas tootmine, antibiootikumi modifitseerimine, alternatiivse
metabolismiraja kasutamine)
Mikroorganismide toitumine ja heksooside katabolism
1. Milliste järgmiste ainete puhul tuleb see aine ATP energia arvelt aktiveerida, et see
saaks siseneda metabolismi?
– Väävel, glükoos
2. Milliste protsesside puhul on elektronide aktseptoriks hapnik?
– aeroobne hingamine, nitrifitseerimine
3. Märgi õiged väited, milliste meetmetega saavad bakterid oma eripinda suurendada.
– väikesed rakumõõtmed, jätked ja filamendid, varuainete terad rakus
4. Märgi millistes protsessides osaleb valdavalt membraane ja millistes substraatne
fosfosüülimine.
– Anaeroobne hingamine - membraanne fosfosüülimine
Aeroobne hingamine - membraanne fosfosüülimine
Fototroofia - membraanne fosfosüülimine
Kääritamine - substraatne fosfosüülimine
Glükolüüs - substraatne fosfosüülimine
5. Milline rada on oluline, et bakterirakud saaks sünteesida lipopolüsahhariide,
aromaatseid aminohappeid ja nukleiinhapete suhkrujääke?
– pentoosfosfaadi rada
6. Reasta heksooside lagundamise põhietapid, kui kasutatakse substraadina
polüsahhariide, mida kasutatakse aeroobselt.
– Monosahhariidide moodustamine
Monosahhariidide lagundamine
Püruvaadi teke glükolüüsis
Atsetüül-CoA teke
Tsitraaditsükkel
Elektrontransportahel
Moodustub CO2 ja vesi
7. Milline neist võis olla esmane energiaallikas ürgsel Maal, sest see tekib ka
elusorganismidest sõltumata keemilistes reaktsioonides
– polüfosfaat
8. Ühenda omavahel energiaallikas, elektronide doonor ja süsinikuallikas ning neile
vastav sobiv toitumistüübi nimetus
– valgus, anorgaaniline aine, CO2 - fotolitoautotroof
keemilised ained, anorgaaniline aine, orgaaniline aine - kemolitoheterotroof
keemilised ained, orgaaniline aine, orgaaniline aine - kemoorganoheterotroof
keemilised ained, anorgaaniline aine, CO2 - kemolitoautotroof
Käärimised
1. Märgi bakterid (või nende rühmad), kes kääritamisel võivad etanooli moodustada.
– heterofermentatiivsed piimhappebakterid, klostriidid, enterobakterid, Zymomonas
mobilis
2. Milliste tunnuste alusel on kääritamistele nimetused antud? Märgi õige
– peamise produkti järgi
3. Mille poolest erinevad homo- ja heterofermentatiivne piimhapekäärimine? Vali
tunnusele vastavad käärimised.
– läbiviijad saavda kasutada substraatset fosforüülimist - homofermentatiivne ja
heterofermentatiivne
toimub kõrgema temperatuuriga keskkonnas - homofermentatiivne
heksoosid kataboliseeritakse glükolüüsis - homofermentatiivne
tekib etanooli - heterofermentatiivne
viivad läbi laktokokid - homofermentatiivne
tekib piimhapet - homofermentatiivne ja heterofermentatiivne
tekib gaasi - heterofermentatiivne
tekib summaarselt 1 ATP - heterofermentatiivne
4. Millises asukohas toimuvad suurima tõenäosusega skeemil
näidatud käärimisprotsessid?
– mäletseja looma vatsas
5. Märgi millist produkti nimetatutest ei moodustu enetrobakterite käärimisel
– butüraat
6. Millise käärimisprotsessiga bakterites on seotud vitamiin B12?
– CO2 fikseerimisel propionaadi tekkega
7. Märgi milline käärimisprodukt on oluline järgmiste ülesannete jaoks:
– piimasuhkru moodustumine lehma maksas - propionaat
šveitsi juustu küpsemine - propionaat
hambakaariese teke - piimhape
õlle valmistamine - etanool
sooleepiteeli tervena hoidmine - butüraat
jogurti valmimine - piimhape
8. Milliste omaduste või tunnuste abil saavad pärmid (Saccharomyces sp) väga
edukalt suhkruid etanooliks kääritada? Vali õiged väited
– palju suhkrute transportereid, taluvad kõrget etanooli konsentratsiooni, kasutavad
osmoprotektorit
Anaeroobne hingamine ja kemolitotroofia
1. Vali, millised nimetatutest on anaeroobsel hingamisel moodustuvad lõppproduktid
ja millised on kemolitotroofsete protsesside lõpp-produktid.
– nitraat - kemolitotroofia
nitrit - anaeroobne hingamine
sulfit - anaeroobne hingamine
vesinik (H2) - anaeroobne hingamine
Fe(III) ühenid - kemolitotroofia
süsihappegaas - kemolitotroofia
sulfaat - kemolitotroofia
suktsinaat - anaeroobne hingamine
2. Millised neist on kemolitotroofsete bakterirühmade
esindajad? Märgi õiged
– rauabakterid, vesinikubakterid, tioonbakterid,
vingugaasibakterid
3. Milline protsess toimub joonisel näidatud hapnikuvabas
tsoonis (anoxic)?
– Väävlihingamine
4. Millised protsessid toimuvad ainult prokarüootide vahendusel? Märgi õiged
– metanogenees, kemolitotroofia, nitrifitseerimine
5. Järjesta elektrontransportahela protsessi komponendid vastavalt elektronide
liikumisele nitraatse hingamise korral.
– dehüdrogenaas (Fp valk)
Fe-S valk
kinoon
tsütokroom b
nitraadi reduktaas
eraldub nitrit
6. Mis on anaeroobne hingamine? Täida lüngad sobivate sõnadega, eralda sõnad
komaga
Anaeroobne hingamine on protsess, kus oksüdeeritavalt substraadilt ärastatud
elektronid kantakse …. erinevale anorgaanilisele elektroni aktseptorile ja ATP
moodustub …. fosforüülimisel
– hapnikust, membraansel
7. Vali bakteriperekond, kes vastavat protsessi läbi viib
– Thiomargarita - väävliühendite oksüdatsioon
Nitrospira - nitrifitseerimine
Desulfovibrio - sulfaatne hingamine
Thiospira - väävliühendite oksüdatsioon
Nitrobacter - nitrifitseerimine
Lämmastiku sidumine ja bakteriaalne fotosüntees
1. Mis on pildil?
– Tsüanobakterid heterotsüstiga
2. Vali, kas nimetatud juuremügaras esinevad komponendid on moodustanud taim või
bakter või mõlemad.
– glutamiin - bakter
sümbiosoomi ümbritsev membraan - taim
leghemoglobiin - taim ja bakter
bakteroidi membraan - bakter
nitrogenaas - bakter
3. Märgi õiged väited lämmastikuringe kohta
– Nii nitraatsel hingamisel kui ka denitrifikatsioonil on mõlemas võtmeensüümiks
nitraadi reduktaas ja elektronide aktseptorina kasutatakse nitraati,
Õhulämmastiku sidumises osalevad nii vabalt elavad kui taimedega sümbioosis olevad
bakterid ja arhed.
4. Vali, kas antud tunnus on iseloomulik purpursetele väävlibakteritele, purpursetele
mitteväävlibakteritele või mõlemale rühmale
– välise redutseerijana võidakse kasutada orgaanilisi ühendeid - purpursed
mitteväävlibakterid
produktina tekib sulfaat - purpursed väävlibakterid
anaeroobid - purpursed väävlibakterid
välise redutseerijana kasutatakse väävelvesinikku, tiosulfaati, rauda - mõlemad
fotolitoautotroofid - purpursed väävlibakterid
5. Järjesta juuremügara tekke etapid alates selle tekke indutseerimisest
– Taimejuured eritavad flavonoide
Mügarbakterid liiguvad juurekarvade lähedusse
Mügarbakterid eritavad nod-falktoreid
Juurekarv kõverdub
Bakterid moodustavad infektsiooniniidi
Bakterid jagunevad
Bakterid tungivad tungivad taimekoesse
Bakterite signaalmolekulide toimel juurekoe rakud jagunevad
Moodustub juuremügar
6. Milline võimalus nimetatutest ei sobi nitrogenaasikompleksi hapniku eest
kaitsmiseks?
– hüdrogenaasi puudumine
7. Reasta Vinogradskiy sambas olevad organismid alates kõige ülemisest kihist
– Tsüanobakterid
Väävli oksüdeerijad
Purpursed mitteväävlibakterid
Purpursed väävlibakterid
Rohelised väävlibakterid
Sulfaatsed hingajad
8. Vali, kas antud organsimirühmad on oksügeensed või anoksügeensed
fotosünteesijad
– tsüanobakterid - oksügeensed
vetikad - oksügeensed
rohelised väävlibakterid - anoksügeensed
õistaimed - oksügeensed
rohelised mitteväävlibakterid - anoksügeensed
purpursed mitteväävlibakterid - anoksügeensed
purpursed väävlibakterid - anoksügeensed
Normaalse mikrobioota funktsioonid inimese jaoks ja faktorid, millest selle koostis sõltub.
Kes on gnotobiondid?
Mikroobivabade loomade eripärad.
Inimese erinevate kehapiirkondade (naha, mao, suuõõne, soolestiku) mikrobioota tunnused ja selle põhirollid.
Sarnased õppematerjalid
![Mikrobio II eksamiks kordamine]()
35
docx
Mikrobio II eksamiks kordamine
1
Teemad kordamiseks 2012
dotsent Tiina Alamäe
Mikroorganismide toitumine. Mikroobide eripind ja kuju, nende seos toitumisega.
Toitumisprobleemid väga suurtel bakteritel. Võimalused eripinna suurendamiseks. Pelagibacter
ubique.
Mikroorganismid toituvad osmootselt kasutavad lahustunud aineid, mis jõuavad nende rakku läbi pinna,
läbides kapsli, kesta ja membraani. Peamiseks takistuseks on rakumembraan, mida ained läbivad kas
difusiooniga või kanaleid ja valgulisi transportereid kasutades. GN bakteritel tuleb täiendava barjäärina
juurde rakukesta välismembraan. Seetõttu on GN bakterid vähem tundlikud mürgistele ainetele. Sh
aintibiotsidele.
Mida väiksemate mõõtmetega bakter, seda suurem eripind. Väikeste mõõtmete tõttu on palju
toitumispinda (suur eripind). Ülilihtsad organismid ei saakski olla väga suured, sest suurena nad ei
toimiks: nad ei suudaks rakku varustada toitainete
![Metabolismi üldiseloomustus]()
15
doc
Metabolismi üldiseloomustus
AINEVAHETUSE ÜLDISELOOMUSTUS
METABOLISMI PÕHIMÕISTED
Metabolism = ainevahetus kõigi elusrakus kulgevate keemiliste reaktsioonide võrk
Katabolism keerulise ehitusega ühendite lagundamisega (degradatsiooniga) seotud reaktsioonide
kogum
Anabolism raku makromolekulide sünteesiga seotud reaktsioonide kogum
Vahemetabolism ainevahetusreaktsioonid, milles osalevad (intermediaarne metabolism) väikesed
molekulid (nn. intermediaadid)
Metaboliidid raku ainevahetuses osalevad ained
Metaboolsed rajad järjestikuste ensüüm reaktsioonide ahelad; ühe lõppprodukt on substraadiks
järgmises reaktsioonis
Metaboolsed rajad on paljuastmelised
· Lineaarsed
· Hargnenud
· Tsüklilised
METABOLISM KULGEB ÜKSIKUTE, KONTROLLITUD
ASTMETENA
Glükoosi kontrollimatul lagundamisel vabaneks korraga suur hulk energiat. Paljuastmelises
ensümaatilises protsessis on vabanevad energiahulgad väikesed (mitte üle 60 kJ/mol) ja ülekanne
individuaalsete
![Bakterite osa küsimused ja vastused]()
6
rtf
Bakterite osa küsimused ja vastused
Kordamisküsimused ja vastused - bakterid ja arhed
Veekogude elustik
1. Millest toituvad bakterid, milliseid elemente vajavad?
Vastavalt süsinikuallikale (metaboolsete protsesside järgi) jagatakse bakterid heterotroofideks ja
autotroofideks. Heterotroofide süsinikuallikaks on orgaanilised ühendid. Autotroofide
süsinikuallikas on süsihappegaas (CO2).
2. Millised on bakterite ja arhede toitumistüübid? Nimeta ja kirjelda lühidalt.
Fotolitotroofid: Taimed, vetikad, tsüanobakterid: valguseenergia arvel sünteesivad ATPd, C-allikana
kasutavad CO2, CO2 redutseerimiseks kasutavad vett. Purpursed ja rohelised väävlibakterid: CO2
redutseerivad H2S abil
Fotoorganotroofid: Purpursed ja rohelised mitteväävlibakterid, ATP sünteesivad valguseenergia
arvel, C-allikana saavad kasutada nii CO2 kui ka orgaanilisi ühendeid, CO2 fikseerimiseks
kasutavad reduktiivjõu allikana enamasti orgaanilisi aineid.
Kemolitotroofid: Nitrifitseeri
![Biokeemia II EKSAMiks kordamine]()
32
docx
Biokeemia II EKSAMiks kordamine
Aminohapete biosüntees
1. Defineerige mis on lämmastiku fikseerimine ja millised organismid on võimelised seda protsessi
läbi viima. Kirjeldage milline on lämmastiku tsükli üldskeem looduses ja millisel kujul on meie
organism võimeline lämmastikku kasutama biosünteetilistes protsessides.
Molekulaarne lämmastik N2 muundatakse redutseeritud või oksüdeeritud vormiks. Atmosfääris leiduv N 2
on keemiliselt väga inertne ning metabolismis kasutamiseks tuleb see redutseerida NH 3 kujule. Toimub UV
kiirguse ja välgu kaasabil maa atmosfääris. Eluslooduses on lämmastikku fikseerima võimelised vähesed
mikroorganismid, kes redutseerivad elementaarse lämmastiku ammooniumiks. Mõned sellistest bakteritest
on vabalt elavad, paljud on aga taimede, eelkõige liblikõieliste taimede, sümbiondid. Valdav enamus
organisme on võimeline omastama lämmastikku NH 4+ vormis.
Summaarne reaktsioon
N2 + 10H+ + 8e- + 16ATP Z 2NH4+ + 16ADP + 16 Pi + H2
![Bioloogia konspekt-aine- ja energiavahetus-ATP-fotosüntees]()
22
docx
Bioloogia konspekt: aine- ja energiavahetus, ATP, fotosüntees
Aine- ja energiavahetus
1. Defineeri mõiste
Autotroofid- enamik organismidest, kes kasutavad energia saamiseks
valgusenergiat ja orgaanilisi aineid toodavad väliskeskkonnast saadud
anorgaanilistest süsinikuühenditest
Heterotroofid- organismis, kes kasutavad energia saamiseks teiste
organismide elutegevuse käigus tekkinud orgaanilisi ühendeid ja
toiduga saadud orgaanilistest ühenditest
Miksotroofid- organismid, kes vastavalt tingimustele võivad olla
valguse käes autotroofid, pimeduses heterotroofid
Rakuhingamine- glükoosi lõplik lagundamine hapniku abil, mille
tulemusena vabanev energia salvestatakse makroergilistesse
ühenditesse (ATP) ja eraldub CO2 ja H20
Makroergilised ühendid- väikesed org. ühendid, mis osalevad
keemilise energia salvestajate ja ülekandjate organismides toimuvates
reaktsioonides
ATP- (adenosiintrifosfaat) peamine rakkudes kasu
![Aeroobsete ja Anaeroobsete bakterite metabolismi erinevused]()
9
docx
Aeroobsete ja Anaeroobsete bakterite metabolismi erinevused
Tartu Ülikool
Arstiteaduskond
Tartu Ülikooli mikrobioloogia instituut
Aeroobsete ja anaeroobsete bakterite
metabolismi erinevused
Referaat
Autor: Eliys Tomson
Arstiteadus II, 7. Rühm
Sõnade arv 1493
![Bioloogia küsimused 11-klassile]()
5
docx
Bioloogia küsimused 11. klassile
Bakterite ehitus
Bakterid on üherakulised eeltuumsed ehk prokarüootsed organismid, mis paljunevad
pooldumisel. Arhed ehk ürgbakterid on rakutuumata ja rakutuumaga rakkude vahepealsed.
Elavad äärmuslikes keskkonna tingimustes.
Ekstermofiilid kuumavee allikad ja ülissoolased järved. Anton van Leeuwenhoeki avastas
bakterid 17 sajandil. Robert Koch avastas haigusi põhjustavaid baktereid(1843 1910).
L.Pastewr avastas käärimise.
Bakterite ehitus
1) Piilid
Vajalikud bakteriraku kinnitumiseks ning vahel ka geneetilise informatsiooni vahetuseks.
2) Kapsel
Ei esine kõikidel bakteritel. Elusorgansmides on kihn oluline kaitsebärjäär, et seista loomse
organismi kaitse mehhanismide vastu.
3) Tsütoplasma ja ribosoomid
Läbi paistev, poolvedel ja sisaldab vett, valge ja rasvu
Ribosoomid:
Sisaldavad RND-d ja proteine; valgusünteesi toimumispaik; Asenduvad mitokondreid
4) Nukleoid ja plasmiidid
Rõngakujuline kromosoom, kuhu on ko
![Biokeemia eksami variandid]()
18
docx
Biokeemia eksami variandid
EKSAMI VARIANDID
I VARIANT
1. Iseloomustage DNA ahela ehitust millistest komponentidest ahel koosneb, millised
kovalentsed sidemed on komponentide vahel ja millised sidemed on ahela ehituslikuks
aluseks
DNA koosneb kahest nukleiinhappe ahelast moodustades kaksikspiraal, milles suhkur-
fosfaat selgroog on väljaspool ja lämmastikalused asuvad heeliksi sisemuses.
Lämmastikalused paarduvad omavahel vesinisidemete abil. Paarid moodustuvad
puriinide ja pürimidiinide vahel.
Nukleiinhappe ahela ehituslikuks aluseks on 3´5´-fosfordiesterside.
2. Kirjutage ensüümireaktsiooni algkiiruse võrrand (Michaelis-Menten'I võrrand) ja
iseloomustage selles olevaid tegureid. Arvutage, millega võrdub suhe v/Vmax, kui
substraadi kontsentratsion ületab 8-kordselt Km väärtust.
v= Kui [S] = Km, siis v = Vmax/ 2.
o Vmax = k2 [ET], (M s-1)
o Km= , (M)
· Vmax on ensüümi iseloomustav konstant
· Vmax on teoreetiline maksimaalne
Meedia
Kommentaarid (0)




































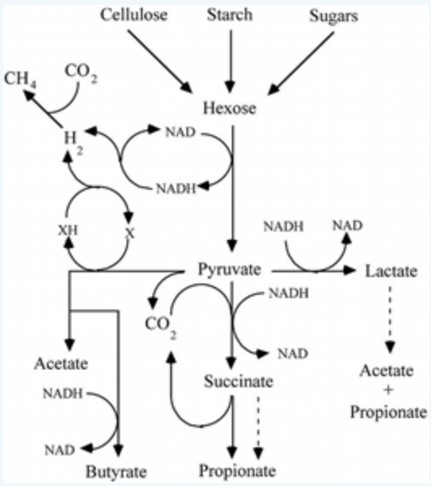
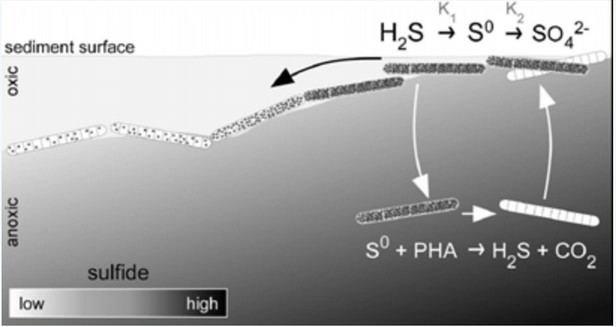
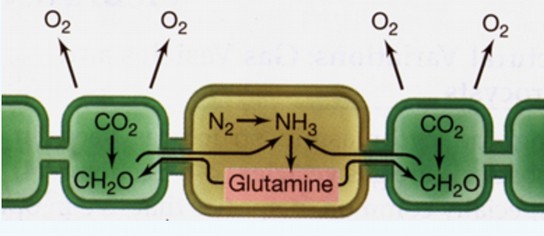







Kõik kommentaarid