G-valkude aktiveerimisel osalevad GEF-id, G-valkude inaktiveerimisel osalevad GAP-id. 3. Mis retseptoritega seonduvad G-valgud? Miks need retseptorid on olulised? G-valk seoselised retseptorid, GPCRid on kõige arvukamad retseptorid rakupinnal GPCRid. Vähedavad palju erinevaid signaale: hormoonid (glükogeeni ja rasva metabolism), neurotransmitterid, valguskiirgus, lõhnad GPCRe kasutatakse palju närvirakkudes – neuronaalsed signaalirajad Signaaliülekanne II 1. Teada 4 põhitüüpi (8 alatüüpi) signaaliülekande rada, mis viib geeniekspressiooni muutusteni! Teada tuleks, milline signaal aktiveerib raja, mis retseptorit kasutatakse, retseptori eripära, millistele valkudele antakse signaal edasi ning kuidas viiakse signaal tuuma! Retseptor-Seriin/Treoniin – kinaasid Tsütokiinide retseptorid ja JAK/STAT rada
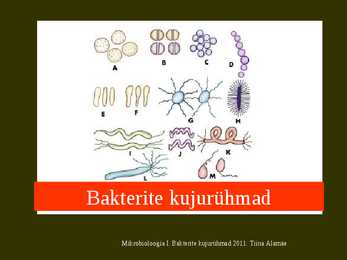
neist nöörduvate spooridega (koniididega). Mõnedel aktinomütseetidel ümbritseb spoore kate esineb sporangium. Mõnede aktinomütseetide spooridel on viburid ja nad saavad vees liikuda. Mikrobioloogia I. Bakterite kujurühmad 2011. T Streptomyces coelicolor'i elutsükkel Koniid (spoor) idaneb ja sellest kasvab välja hüüf, mis pikeneb otsast. Moodustub substraadimütseel. Kui toitainete sisaldus keskkonnas langeb, siis aktiveeruvad signaalirajad, mis algatavad õhumütseeli moodustumise. Õhumütseeli hüüfide tipu fragmenteerudes moodustuvad koniidid. Koniidid on paljunemisvahenditeks. Mikrobioloogia I. Bakterite kujurühmad 2011. Tiina Alamäe Koniidid moodustuvad õhumütseeli hüüfitippude fragmenteerudes. Nad on hüdrofoobse pinnaga, kerged ja kanduvad tuulega laiali. Nende idanedes saab alguse uus aktinomütseedi koloonia. Mikrobioloogia I. Bakterite kujurühmad 2011
ajavad oma "eetika" teemat. 15. Missugused oleksid potentsiaalsed, rakuteraapias kasutatavad tüvirakkude allikad? Blastotsüst, mis on saadud munarakust, kust on eemaldatud tuum ning sisestatud somaatiline rakutuum; indutseeritud tüvirakud; loote aju, üldse närvikude; neuraalharjarakud, skeletilihase (vöötlihase) tüvirakud, epiteelirakkude eellased pärisnahas ja seedesüsteemis, kõhunäärme ja maksa tüvirakud. 16. Arengulised signaalirajad WNT beeta-kateniin on tsütoskeleti komponent, mis translokeerub tuuma, kus funktsioneerib kui TF; WNT'i sidumisel Frizzled retseptorile inhibeerib DSH GSK-3b aktiivsuse. Fosforüülitud beeta-kateniin ubikvinteeritakse ja lagundatakse proteosoomides. Mittefosforüülitud beeta-kateniin on stabiilne ja translotseerub tuuma, kus funktsioneerib koos TCFga kui TF. Beeta-kateniini/TCF kompleks aktiveerib geene, mis osalevad rakkude elushoidmises, proliferatsioonis ja arengulises
efektor-rakkudeks. Th rakus aktiveeritakse mitmed geenid, millede produktid on olulised interaktsioonides AG-ga ja äratundmisel. 1) vahetud (ekspresseeritakse 30' peale AG äratundmist); kodeerivad transkriptsioonifaktoreid c-Fos, c-Myc, c-Jun, NF-AT, NF-kB. 2) varajased (1-2h jooksul); kodeerivad erinevaid tsütokiine: IL-2; IL-3; IL-6; IFN jt). 3) hilised geenid ekspresseeritakse enam kui 2p hiljem st.peale AG tundmist; kodeerivad adhesioonimolekule. Aktiveeritakse erinevad signaalirajad. 20. T lümfotsüütide küpsemine harknäärmes e. tüünuses Tüvirakud pärinevad luuüdist, nende pinnaretseptorid määravad ära migreerumise tüümusesse. Tüümus jõuab max suuruseni puberteedis, seejärel oluliselt väheneb ning edaspidi saab uusi T-rakke: 1) pikaealised T mälurakud; 2) T-rakud valmivad mujal (N:maksas); 3) väike osa tüümusest säilib. Interaktsioonid, kus
Th rakus aktiveeritakse mitmed geenid, millede produktid on olulised interaktsioonides AG-ga ja äratundmisel. 1) vahetud (ekspresseeritakse 30' peale AG äratundmist); kodeerivad transkriptsioonifaktoreid c-Fos, c-Myc, c-Jun, NF-AT, NF-kB. 2) varajased (1-2h jooksul); kodeerivad erinevaid tsütokiine: IL-2; IL-3; IL-6; IFN jt). 3) hilised geenid ekspresseeritakse enam kui 2p hiljem st.peale AG tundmist; kodeerivad adhesioonimolekule. Aktiveeritakse erinevad signaalirajad. BCR ja TCR Kuna mõlemad retseptoritel on väga lühike tsütoplasmaatiline domään, peavad nad assotsieeruma invariantsete signaali ülekande molekulidega, selleks, et genereerida rakusisene signaal. IgIg BCR jaoks CD3 TCR jaoks. BCR on raske ahel ja kappa/lambda ahel. BCR retseptoriks on antikeha. Multivalentsed antigeenid saavad otse seonduda BCR ja sellega põhjustada BCR cross-linki, mis tagab signaali ülekande. TCR on alfa/beeta/gamma/delta ahelad. TCR on heterodimeer
Tsütoplasmaatilised PPR (pattern recognition receptors) NOD –(Nucleotide oligomerization domain) retseptorid . Üle 20 imetajatel (NOD2-muramüül dipeptiid) RLR retseptorid RNA helikaasid Taimede PPR retseptorid(XA21, FLS2) MBL (mannan-binding lectin) C-reaktiivne valk Komplemendi retseptorid RNA helikaasid RIG-I ja MDA5 RIG-I ja akronüümid immunoloogilistes tekstides RIG-I(retinoic-acid-inducible protein ) retseptorite (tsütosoolsed RNA helikaasid) käivitatud signaalirajad Taimede immuunsüsteemi elemente PAMPS- Pathogen-associated molecular pattern LRR- leucine-rich repeat (leutsiini rikaste kordustega retseptorid) R genes (resistentsuse geenid) RNA interference Lihtsamad keemilised mediaatorid ja signaalnolekulid (salitsüül hape,jasmon hape, etüleen.....) Immuunreaktsioonid taimedel miRNA ssRNA molekule mõnest tuhandest kuni 40,000 molekulini raku kohta Leitud kõigis metazoa liikides, 0.5-1% geenidest
ning algab follikulogenees, mille käigus individuaalne arenev munarakk ümbritsetakse folliikuliga. Lisaks osalevad folliikulite tekkes ka sidekoelised teeka rakud, mis on pärit munasarja mesenhümaalsetest eellasrakkudest. Gartner`i juha – Wolffi juha jäänuk naistel Paroöphoron/Epoöphoron – mesonefriliste torukeste jäänukid 91. Milleks on oluline SRY? (mis juhtub kui see geen defektne) Geneetilised faktorid ja signaalirajad gonaadide arengus Enamikul imetajatel on peamiseks bipotentsiaalse gonaadi suunajaks testise arenguteele Y-kromosoomi lühikeses õlas paiknev geen Sry (sex-determining region of the Y chromosme), mis võimendab TF Sox9 ekspressiooni ja läbi selle geenide ekspressiooni, mis on vajalikud testise välja kujunemiseks ja spetsiifilisuse säilitamiseks (inihibeerivad munasarja arengusuunale spetsiifiliste Rspo1, Wnt4, Fβ-perekonna (ingl Transforming growth factor β) liige oxl2 ekspressiooni).
Kahekomponendilistel regulatsiooniradadel on üldiselt signaali ülekandumiseks partnerid konserveerunud. See tähendab, et enamikel juhtudel on sensoril kindel regulaator, mida ta fosforüleerib, kuigi on kirjeldatud ka ristfosforüleerimist (üks sensor fosforüleerib mitut regulaatorit). E. coli'l on kirjeldatud 62 kahekomponendilist regulatsioonirada, Bacillus subtilis'el 70. Varem arvati, et nii paljude signaaliradade ning fosforüleerimiseks konserveerunud aminohapete olemasolul võiks signaalirajad olla suhteliselt kergesti ristreguleeritavad. Seda kinnitasid ka in vitro eksperimendid, kus leiti, et sensor võib fosforüleerida pea 84 igat regulaatorit. In vivo see nii pole ning on teada ainult mõni regulatsioonirada, kus signaal antakse edasi mitmele regulaatorile. Näiteks E. coli kemotaksise sensor CheA fosforüleerib nii CheY-t kui ka CheB-d. Bacillus subtilise Spo0F võib
bakteriaalse infektsiooni vastu võitlemisel. NLRid seovad tsütoplasmas bakteri derivaate, nt peptiidoglükaani. RLR (RIG-1 like receptor) on samuti perekond tsütoplasmaatilisi molekule, mis on olulised viirusinfektsioonide vastu võitlemisel. Äratuntavaks viiruste molekuliks on dsRNA. Osadel RLR retseptoritel on CARD (caspase activation and recruitment domain) domään, mille kaudu aktiveeritakse signaalirajad, mille tulemusena sünteesitakse tüüp I interferoone. Makrofaagide ja dendriitrakkude poolt produtseeritavad tsütokiinid. Nende tähendus immuunsüsteemi funktsioneerimisel. Kaasasündinud immuunsüsteemi rakkude poolt toodetud tsütokiinid aktiveerivad leukotsüüte ja põhjustavad süsteemi ümberkorraldusi; suureneb effektor rakkude süntees ja antimikroobsete valkude süntees. Dendriitrakud produtseerivad tsütokiine IL-12, IL-23 ja IL-27.