Ökoloogia kursuse konspekt. (0)
ÖKOLOOGIA print 1 ilma osade joonisteta
1
ÖKOLOOGIA
- teadus organismide ja keskkonna vahelistest suhetest
- Ernst Haevel 1866
Me mõjutame üksteist vastastikku.
Nt: Parfüümi kandmine – püüd midagi mitte signaliseerida. Püüd keskkonnas paremini
hakkama saada.
Ökoloogia tsentraalne termine: ÖKOLOOGILINE FAKTOR.
• Tegur. Igasugune aine, energia- või infovoog, mis elusorganisme otseselt või kaudselt
mõjutab.
Nt: parfüümi lõhn on infovoog – tajume seda
Assimileerida = enda sarnaseks tegema. Lihtsamatest ainetest keerukamaid organismile
omaseid aineid üles ehitama.
Nt: Mullast saadud mineraalainetest ja süsihappegaasist assimileerib taim orgaanilisi ühendeid.
Akumuleerima = koguma, salvestama.
Nt: Taimed akumuleerivad footonite energiat
ÖKOLOOGILISED FAKTORID:
1. Abiootilised faktorid – eluta faktorid Nt: päike
2. Biootilised faktorid – elus faktorid Nt: Toit
1. Tingimusfaktorid - Tingimusfaktorid teevad meile võimalikuks ressursside
akumuleerimise
Nt: temperatuur, pH, soolsus, infovoog
2. Ressursid
• PAR
• CO2
• H2O
• O2
• Mullamineraalid
• Elusorganismid
Kemosüntees – Orgaanilisi ühendeid saadakse keemilisest energiast (mitte footoni energiast)
Autotroofid – primaarne produktsioon (taimed. Orgaaniliste ühendite süntees anorgaanilistest
ühenditest)
Heterotroofid – sekundaarne produktsioon (loomad ja seened. Toituvad ühenditest, mille on
produtseerinud autotroofid)
Maakoore nihete tulemusena tekivad süvaookeanites kohad, kus magma puutub kokku veega
ja sinna tekivad mustad suitsetajad. Seal on täiesti erakordne ökospsteem. Seal elavad
kemosünteesivad bakterid.
2
ÖKOLOOGIA TASEMED
Organisatsiooni tase eluslooduses viitab mingile hierarhilisele süsteemile.
Ökoloogiat saame uurida erinevatel organisatsioonilistel tasemetel.
1. ORGANI TASE – Ökofüsioloogia
2. ISENDI TASE – Autoökoloogia
2.1 Unitaarne organism – kõik lihtne. Saab selgelt eristada, kust organism algab ja
kust lõpeb.
2.2 Modulaarne organism – keeruline. Organism koosneb moodulitest, mis on
suuremal / vähemal määral iseseisvad.
Nt: maikelluke
Ramet moodustab geneti.
Nt: harilik kuusk. Kuuse oks – 1 tüvest lähtuv oodul. Kui oks end ise ei suuda ülevad pidada
(on varjus/kahjustunud) siis lastakse oksal ära kuivada.
3. POPULATSIOONI TASE – Demograafiline ökoloogia
Isendid samast bioloogilisest liigist moodusavad populatsiooni. Üksi isendid hakkama ei saa,
ja kui saavadki – surevad varsti vlja.
Populatsioon...
1) ... vabalt ristuvate isendite kogum
DEEM – ehk pammiktiline populatsioon
2) ...mingi liigi isendid mingil ajal mingis kohas.
Operatsiooniline populatsioon
Populatsioon võib seega olla ka klassiruum, kus toimub info vahetus, puudub aga vaba
ristumine.
Metapopulatsioon – Omavahel aktiivselt geneetilist materjali vahetav populatsioon.
Nt: erinevad rabad, kus kasvavad jõhvikapopulatsioonid.
Geenid levivad õietolmuga, loomade väljaheidetega
jne.
LEVIS ehk DIASPOOR = organ, mis on mõeldud
ruumis ühest populatsioonist teise liikumiseks.
3
4. KOOSLUSE TASE – Sünökoloogia/Kooslusökoloogia
Koosluse moodustavad omavahel koos elavad populatsioonid
Mõiste „eluskoosulus“ – A.D de la Malle 1825
„societe“ prantsuse keeles kooslus. Tegu on vanima ökoloogia terminiga.
Koosluses elavad populatsioonid mõjutavad üksteist suuremal/vähemal määral.
Kooslus elab kindlas abiootilises keskkonnas.
Lisades kooslusele biootilise keskkonna – saame ökosüsteemi.
5. ÖKOSÜSTEEMI TASE – Süsteemi ökoloogia
Ökosüsteem = kooslus + eluta keskkond - A.Tansley 1935
Süsteemi ökoloogias vaadatakse koos kõikvõimalikke aine/info jne voogusid, mis võivad
organismi mõjutada.
BIOOM – Sarnaste ökosüsteemide kogum üle terve maailma.
„Bioom“ tähistab mingit sorti ökosüsteemi.
Nt: tundra bioom
Nt: meie elame 2 bioomi segus (piiril). Taiga ja nemoraalne (kõigi Eesti metsade)
segabioomis – boreonemoraalne bioom
BIOSFÄÄR – Kõik maakera bioomid kokku.
Biosfäär = globaalne ökosüsteem.
KOKKUVÕTE:
1. ORGANI TASE – Ökofüsioloogia
2. ISENDI TASE – Autoökoloogia
3. POPULATSIOONI TASE – Demograafiline ökoloogia
4. KOOSLUSE TASE – Sünökoloogia/Kooslusökoloogia
5. ÖKOSÜSTEEMI TASE – Süsteemi ökoloogia
4
RESSURSIFAKTORID
1. PAR
PAR paneb aluse ökosüsteemides energiaringele.
PAR – kindlate omadustega kiirgus.
(UV) (violetne) – 380-710 nm – (FR) (infrapuna, punane)
PARi kohta pole vale öelda lihtalt valgus, sest antud lainevahemikku jääb inimesele silmaga
nähtav valgus.
Infrapuna neelab atmosfäär selektiivselt: 7-8%
UV-d neelab atmosfäär enim: umbes 50%
Fotosünteesi aparaadi keskmeks on klorofüll.
Punase ja Sinise valgusega töötab klorofüll kõige
intensiivsemalt.
Rohelist valgust peegeldub enim – sellepärast paistavad
taimed meile rohelisena
Klorofüll on võimeline ergastunud elektrone muutma keemiliseks energiaks. Toimub vee
fotolüüs.
CO2 on madala energiaga aine. Suhkur on aga väga energiarikas.
Klorofülli aitavad lisapigmendid, kromoplastid, mis jagunevad kahte rühma:
1) Kartonioidid – neelavad hästi kollast/punast valgust
2) Ksantofüllid
Nad akumuleerivad karotinoid-pigmente, millest sõltub paljude taimede õite kollane, oranž või
punane värvus. Meelitavad kohale tolmeldajaid ja viljade levitajaid.
Nad on võimelised ergastunud elektrone klorofüllile edasi andma.
UV valgust ega FR valgust ükski pigment neelata ei suuda!
Fotoautotroofid – saabad energia päikeselt
Kemoautotroofid – kasutavad keemilise energia allikaid
5
PARi globaalne kättesaadavus varieerub
Eesti – 0.84
Ekvaatoril – 2.1
PARi intensiivsus varieerub kuni 2 korda!!
Samal laiuskraadil:
maismaal 1.68 (kõrbes)
vees 2.1
Liivatormide tõttu on atmosfäär kõrbete kohal veidi udune ja seetõttu ja pilvisem, sest pilve
moodustamiseks on vaja tolmukübe, mille ümber moodustub tilk.
Mida madalamal on päike, seda vähem energiat pinnaühiku kohta maale jõuab.
Kui palju on PARi langenud?
PARi aastane jaotus Hollandis ja ekvatoriaal-Aafrikas
Ekvaatoril tolm hägustab atmosfääri.
Eestis on taimed vähesege valgusega kohastunud – harjunud pika puhkeperioodiga.
Fotosünteesi efektiivsus
1) Päikesevalgus ei taba fotosünteesi aparaati. Annab
energia ära ja peegeldub infrapunakiirgusena tagasi
atmosfääri.
2) Päikesevalgus ei taba fotosünteesi aparaati ja valgus
läheb läbi lehe, maapinnale
3) Valgus tabab fotosünteesi aparaati ja kvandi energia
kasutatakse fotosünteesiks. Päikesevalgusest saadud energia
abil pannakse süsiniku külge H aatomid.
6
Fotosünteesi summaarne efektiivsus = kui suur hulk taimkattele langevast PARist kasutatakse
ära fotosünteesiks.
Fotosünteesi summaarne efektiivsus on alla 3%.
Vihmametsades 1,2 %
Eesti segametsas 0,6%
PARi jaotumine
Kuuse segamets
Metsavööndi väiksemad taimed elavad pimeduses.
HÄIL – metsalagendik, mille on tekitanud ümberkukkunud puu.
Siis saavad kasvuhoo sisse taimed, kelleni muidu valgus väga ei
jõua
Männik vaesel mullal
Päevalillepõld
Maisipõld
7
Fotosünteesi kõver
Mida rohkem on taimedel valgust, seda kiiremini nad kasvavad.
On taimi, mis vajavad rohkem valgust ja taimi, mis vajavad vähem.
Fotoinhibitsioon – fotosüntees läheb kinni liigse valguse korral.
Fotosünteesi kompensatsioonipunkt – tähistab minimaalset võimalikku PARi taset, mille
juures roheline taim suudab rahuldada oma energiavajaduse. Nii pime, kuidas taim suudab ellu
jääda.
Mida pimedamas on taim kohastunud elama, seda madalam on tema kompenstsioonipunkt.
8
2. CO2 (ja fotosüntees)
• Tähtsaim süsinikuallikas.
Enamus fotosünteesist toimub maismaal.
Maismaal: CO2 on maismaa ainuke süsinikuallikas
Ookeanis: lisaks CO2 on süsinikuallikaks ka HCO3 ja CO32-
Süsinikuallikate intensiivsus:
Seega: mida aluselisem on keskkond, seda raskem on fotosüntees. Aluseline keskkond
raskendab fotosünteesi.
Atmosfääri CO2 sisaldus
Fotosüntees tekkis, kui tekkisid sinivetikad ja 1
tsüanobakter kolis primaarse endotsütoosi teel teise
organismi elama ja temast sai kloroplast.
Sellest ajast saati on fotosünteesi aparaat säilunud
originaalsel kujul.
Tänapäeval elavad taimed pidevas süsinikunäljas.
CO2 on lisandunud 60 aasta jooksul umbes
kolmandiku võrra.
Graafiku CO2 üles/alla kõikumised – aastaaegadest
põhjustatud:
Kevadel – CO2 sisaldus tõuseb
Sügisel – CO2 sisaldus langeb
(lõunapoolkeral vastupidi)
Miks on äkiline süsihappegaasi tõus halb kui kunagi
oli selle konsentsatsioon kordades suurem?
Süsiniku konsentsatsiooni tõus on toimunud enneolematult kiiresti ja loodus ei jõua
sellega kohastuda.
Kasvuhooneefekt – CO2 on 1 kasvuhoonegaasidest.
CO2
HCO3
CO32-
Süsiniku hulk sõltub pH-st:
pH (happeline) – valdavalt lahustunud CO2
pH (aluseline) – kaldub HCO3 ja CO32- poole
9
Kasvuhooneefekt
Kasvuhoonegaasid: H2O, CH4, CO2, NOx, N2O
Kasvuhoonegaasid lasevad läbi PARi, aga neelavad FRi. Nii jääb
energia kasvuhoone sisse.
Positiivse tagasiside põhimõttel:
1. Sulab igikelts (tundrad püsivalt külmunud, mikroorganismide
rikkad)
2. Anaeroobseb bakterid hakkavad tegelema, meeletu N2O ja CH4 pump atmosfääri
3. Veel suurem hulk kasvuhoonegaase.
Mida rohkem on, seda rohkem tekib.
Fotosüntees
6CO2 + H2O + e =(UV/klorofüll/OR)= C6H12O6 + 6O2
OR – oksüdatiivne respiratsioon.
Taimed elavad mineraalse süsiniku näljas.
Fotorespiratsioon
Kui CO2 on vähe, siis paneb valk RnBisCo (äärmiselt ebaefektiivne katalüsaator) fotosünteesi
tsükli tagurpidi tööle. Algab valgushingamine ning see on mõttetu raiskav protsess.
Kui kuuma ja kuivuse tulemusena õhulõhed sulguvad veepuuduse tekke vältimiseks, siis
fotosüntees taimes toimub edasi. Nii ei pääse tekkiv O2 välja ja sisse ei pääse vajalik CO2. Siis
lülitub O2 CO2 asemel Calvini tsüklisse ja algab fosforespiratsioon, kus lõpp-produktideks on
CO2 ja H2O.
Kui taim avab oma õhulõhe, hakkab ta koheselt kaotama ka
vett.
Selleks, et vältida valgushingamist peab taim CO2 juurde
saama, aga õhulõhe pikaajalisel lahti hoitmisel kaotab ta
vett. Ariidses kliimas õhulõhede pikalt lahti hoidmine väga
halb valik.
Taimed jagunevad vastavalt fotosünteesi tüüpidele kolmeks:
1. C3 – taimed: umbes 85% taimedest. Tavaline fotosüntees.
CO2 seotakse läbi Calvini tsükli, mis toimub vaid mesofülli
rakkudes.
(Mesofülli rakud asuvad taime lehtedes või varte erikoores ning
neis on palju kloroplaste kus toimub fotosüntees).
Eestis: pea kõik taimed
10
2. C4 – taimed: kaks protsessi on eraldatud füüsiliselt ja ruumiliselt.
Nad ei liida CO2 Calvini tsüklisse otse õhust. Kui C3 taimedes
fikseeritakse süsinik ainult mesofüülirakkudes, siis C4 taimedes
on see jagunenud mesofüüli- ja juhtkimbu rakkude vahel.
Esmane CO2 sidumine toimub mesofüüli rakkudes (nagu ka C3
taimedes). Seal aga paneb ensüüm PEP karboksülaas CO2 PEP
molekuli külge. Tulemuseks on neljasüsinikuline oksaaloatsetaat
(siit tuleb C4 nimetus).
Seega ei liideta CO2 Calvini tsüklisse otse õhust.
Oksaloatsetaat + NADH + H+ = Malaat + NAD+
PEP karboksülaas – ensüüm, mis on võimeline siduma
elektrone.
Nii transporditakse malaat juhtsoonte küljes asuvatesse kimbuümbrisrakkudesse, millel
puuduvad õhulõhed. Seal toimub süsiniku sidumine ja Calvini tsükkel
Lühidalt: kaks protsessi on omavahel füüsiliselt ja ruumiliselt eraldatud:
1. Süsiniku sidumine
2. Calvini tsükkel
Eelis: Calvini tsükkel on pidevalt CO2 varustatud, ka siis, kui õhulõhed on suletud.
C3 ja C4 taimede erinevus seisneb süsihappegaasi sidumises.
Eestis: raud-soodahein ja muidu nt mais
3. CAM – taimed: Ainult väga kuumades kuivades kohtades kasvavad paksulehelised timed.
Toimub happe-metabolism.
Süsinik fikseerub ööpäeva tsükli alusel – fotosünteesietappide
jagamine aitab hoida kokku vett.
Öösel – õhulõhed avanevad ööjaheduse saabudes kui õhuniiskus
tõuseb ja algab CO2 sidumine – minimaalne veekadu.
Päeval – Õhulõhed sulguvad päeva saabudes. Fotosünteesiks
hakatakse
vakuoolidesse
talletatud
malaadist
eraldama
süsihappegaadi, mis sisanab Calvini tsüklisse.
Eestis: kukehari
Calvini tsükkel
11
3. Hapnik
Enne elu tekkimist maal vaba hapnikku ei eksisteerinud.
Raud ei olnud maal oksüdeeritud kujul, enamus oli Püriit – FeS2 . Püriit oksüdeerub hapniku
olemasolul väga kiiresti.
Vaba hapniku tekkimisel:
1. 58% oksüdeerus rauaga (Fe2O3)
2. 38% reageeris väävliga (SO42-)
3. 4% on jäänud üle. 4% on molekulaarse hapniku hulk tänapäeval atmosfääris.
20.9% - tänapäeva molekulaarne hapniku kontsentratsioon atmosfääris.
Maakoores peaks iga kunagise hapniku aatomi kohta olema 1 redutseeritud süsiniku aatom.
Kus on – ei tea :)
Hapnik veekogudes
Talvel - jääkihi all hapniku hulk aina väheneb, aga juurde
ei tule.
Süvaookeanites - hapnik tuleb ainult ülemistest
veekihtidest, kuhu hapnik lahustub.
Hapniku kättesaadavus veest on sõltuvuses
temperatuuriga:
Mida madalam temperatuur, seda rohkem hapnikku kätte
saab.
Soojad ja seisvad veed - hapnikuvaegsed
Hapniku kontsentratsioon maal läbi aja
2 miljardit aastat tagasi – võrreldes praegusega atmosfääris 0,01%
1,4 miljardit aastat tagasi – võrreldes praegusega atmosfääris 2%
560 miljonit aastat tagasi = kambriumi plahvatus – võrreldes praegusega atmosfääris 10%
Hapniku kumulatiivne produktsioon fotosünteesijate poolt alates elu tekkimisest ca 4 miljardit
aastat tagasi.
12
4. Vesi
Suur pall – kogu maakera vesi
läbimõõt 1384km
Väike pall – kogu maakera magevesi
läbimõõt 273km
Imepisike pall – jõgede ja järvede magevesi
läbimõõt 56km
5. Keemilised elemendid
Taimed saavad keemilised elemendid algselt mullast.
Na ja Cr – vajalikud ainult loomadele.
Kloriidid – taimedele kahjulikud.
K – vajalik vaid õistaimedele.
N ja P – Olulisimad vees, eriti maismaaveekogudes elavatele organismidele
N: Kõik lämmastikku sisaldavad soolad dissotseeruvad lihtsalt, leostuvad vesilahuses mullast
välja. Taimed elavad sümbioosis bakteritega, kes suudavad lämmastikku siduda.
P: Fosfor on küllaltki kapriisne, annab teatud tingimustel liikumatuid ühendeid. See sõltub
mulla reaktsioonist.
Mullal neutraalne pH – kõik on ok
Mullal neutraalselt kõrgem/madalam pH – tekivad liikumatud kompleksid, mis pole
taimedele kättesaadavad.
Suuremas koguses on kõigil elusorganismidel vaja makroelemente, Mikroelemente on vaja
väiksemates kogustes (kuid on sama olulised).
Soodes ja süvaookeanites– vesi on anoksiline. Hapnik ei difundeeru seisvasse vette.
Seal on oksüdatiivne respiratsioon tugevasti häiritud.
Soodes on turvas – lagunemata taimejäänused.
Suur pall – kogu maakera vesi
(läbimõõt 1384 km);
Väike pall – kogu maakera
magevesi (läbimõõt 273 km);
Imepisike pall – jõgede ja
järvede magevesi (läbimõõt 56
km).
13
KOKKUVÕTE RESSURSIFAKTORITEST:
1. PAR
2. CO2
3. Hapnik
4. Vesi
5. Keemilised elemendid.
14
TINGIMUSFAKTORID
1. Temperatuur
Vesi läheb tahkeks küllaltki kõrgetel temperatuuridel. Jääs aktiivset elutegevust toimuda ei saa.
Veel on suur soojusmahtuvus! Vesi mahutab palju sooja. Suur soojusmahtuvus ka annab palju
sooja.
Ookeanid: puhverdavad temperatuurikõikumisi. Kontinentaalses kliimas on suvised/talvised
kõikumised drastiliselt suuremad.
Ida-Siberis on talve ja suve temperatuuride vahe 100 kraadi.
Ehk: vesi tekitab elatava temperatuuri.
Maakera on üpris jahe, sest 71% maakerast moodustavad ookeanid, mille keskmine
temperatuur on 2 kraadi (süvaookeanid ja jää ja liustikud kaasa arvatud)
Maakera keskmine temperatuur linnas: 14.7 kraadi
Maakera keskmine temperatuur on viimase 35 aastaga tõusnud 0.75 kraadi. Aina
rohkem PARi jääb maakerale kinni.
Jääaeg = kui on olemas mandrijää kilbid. Praegu on jääaeg.
Registreeritud ekstreemsed temperatuurid:
Maksimum: California Death Valley 56.7 kraadi
Miinimum: Antarktikas -89.2 kraadi
Temperatuur on seotud laiuskraadiga ja kõrgusega
Kehtib seaduspära:
100m kõrgust = -1 kraadi aastast temperatuuri kuivas õhus
= -0.5 kraadi aastast temperatuuri niiskes õhus.
Sõltuvus laiuskraadidest:
• Maakera telg on kaldu
• Maakera trajektoor on elliptiline
Apheelia – maa asetsemine päikesest kõige kaugemal (juuli)
Periheelia – maa asetsemine päikesele kõige lähemal. Maa saab 6% rohkem kiirgust. (jaanuar)
Seetõttu pole meil talv nii külm kui võiks ja suvi nii soe kui võiks.
Organismid on kohastunud temperatuurikõikumisi trotsima erinevalt
1. POIKILOTERMSED = kõigusõõjased organismid.
Organismidel puudub hea meede temperatuuri reguleerimiseks.
2. HOMEOTERMSED = püsisoojased organismid.
Ainult imetajad ja linnud. Evolutsiooni tippsaavutus. Homeotermsed organismid suudavad
piisavalt energiarikka toidu olemasolul trotsida temperatuurikõikumisi.
Nt: keiserpingviinid. Isased hauduvad mune pool aastat ja nad ei söö see periood.
15
3. HETEROTERMSED – Suudavad püsisoojalisusest loobuda ebasoodsate tingimuste
üle
Elamiseks.
Nt: kõik loomad, kes magavad talveund. Siis on füsioloogiliselt mõistlik viia kehatemperatuur
nii madalale kui võimalik.
Nt: nahkhiired ja pingviinid. Pingviinide jalad on külmad, et nad ei sulataks lund ja ei tekiks
jääkamakaid jalgade ümber.
KONN: POIKOTERMILISE LOOMA MUDEL.
Konna kehatemperatuuri mõjutavad 9 ökoloogilist, teineteisest sõltumatut faktorit.
Soojus tuleb:
1. Hajus päikesekiirgus
2. Otsene päikerekiirgus
3. Peegeldunud päikesekiirgus
Konna jahutavad:
4. Tagasikiirgus
5. Evaporatsioon
Tuleb või läheb, vastavalt temperatuuride erinevusele:
6. Konvektsioonne soojusvahetus – Temperatuurid püüavad ühtlustuda, TD II seadus
7. Soojusvahetus teiste objektidega
8. Soojusvahetud maapinnaga
9. Soojusvahetus teiste organismidega – Karjas on kiirgusvahetus väga oluline.
16
Organismide paljunemise sõltuvus temperatuurist
Temperatuurist sõltub ontogenees – organismi arenguks kuluv aeg.
Mida soojem temperatuur, seda kiirem areng.
Positiivne temperatuur taimedele on üle 5 kraadi.
See mõõdetakse kraadpäevade arvuga
Nt: t=7C siis see on +2C
Ja kui temperatuur 7 kraadi kestab 10 päeva: 2*10=20 kraadpäeva.
Temperatuurialaste rekordite hoidjad:
Tedostromia ablangifolia (death Valleyst) – fotosüneesib 50 kraadi juures
Bakter Pyrodictumaccultur – optimum on 105 kraadi.
Külmadel temperatuuridel piire pole.
Siberi Lehise kaitsemehhanism külmumise vastu: Rakkude sees olev lahus on hästi
kontsentreeritud, mis alandab külmumistemperatuuri. Rakkudevaheline lahus on lahja ning
see jäätud, rakud seest ei jäätu.
Aasta keskmine sademete hulk on kooskõlas temperatuuriga!
Väga külm = väga kuiv
Väga soe = väga märg
Külmas lasevad taimed lehed maha – lehtedest aurab vett.
Maakera keskmise aastase temperatuuri muutumine:
Ühe rohutirtsu ja ühe liblikavastse areng
Sõltuvalt temperatuurist
17
2. pH
elu on pH vahemikus: 3,5 – 9
Muld – maapinna pealmine kobe kiht, mida on oluliselt ümber kujundanud taimed ja
mikroobid.
pH mullavees, on 3-10
Mulla pH sõltub peamiselt keemilisest koostusest ja sademete hulgast.
Keemilises koostises on kesksel kohal karbonaadid. Põhiliselt magneesium- ja
kaltsiumkarbonaadid MgCO3 ja CaCO3. Karbonaadid annavad mullale aluselisust.
Kui karbonaate on mullas vähe, uhub vesi vähesedki karbonaadid mullast minema ja
alles jäävad silikaadid ja muud happelised soolad.
Eestis: happelised (leostunud) mullad + veidi neutraalseid muldasid
Põhja-Aafrikas: aluselised mullad
Miks on mulla pH tähtis?
Mulla pH mõjutab oluliste mullamineraalide ja toiteelementide kättesaadavust – mõjutab nende
lahustuvust ja seeläbi kättesaadavust.
Fosfor (P): Hästi kättesaadav pH 5,5-7,5. Fosfor hakkab
piirama produktsiooni alustelistes ja happelistes
muldades.
Alumiinium (Al): Liikuv vaid happelistes muldades.
Alates pH 6 on toksiline!! Taimedel on selle vastu oma
kohastumused
pH rekordite hoidjad:
arhe Sulfolobus ac.daldaru: pH opt = 2 (+90 kraadi)
Soodajärvede soodabakter pH opt = 13
:) Pandadel kulub pool assimileeritud energiast toksiinide kahjutuks tegemiseks.
Mulla
pH
globaalne jaotus
punane: happeline
Kollane:
neutraalne
Sinine: aluseline
Must: ei tea
Mulla pH globaalne jaotus (punane – happeline muld; kollane –
neutraalne muld; sinine – aluseline muld; must – andmed puuduvad)
18
3. Soolsus
Soolsus on tingimusfaktor, sest taimedel tuleb kõik üleliigsed soolad tõrjuda. Maismaamullas
on suur soolsus ja enamus taimed seda ei talu. Taimed, kes suurt soolsust taluvad, omavad
selleks vastavaid kohastumusi.
Peamisteks sooladeks mullas: MgCl, MgSO4, KNO3, NaHCO3.
Ookeani keskmine soolsus on 35‰.
Madalam soolsus: arkliline ookean
Kohati soolased paigad: atlandi ookean
Soolasem: vahemeri
Ookeani soolsuse globaalne jaotus
Vee jaotumine vastavalt soolsusele:
Soolsus > 0,5 ‰ – magevesi
Soolsus 0,5 – 30‰ – riinvesi
Peamiselt laguunides ja kitsa suudmega meredes, kes
saavad oma vee põhiliselt järvedest. Läänemeri.
Läänemerre avanevad paljud jõed, kuid soolast vett
lisandub vaid läbi Taani väina. Nii saavad läänemeres
hakkama osad magevee kalad ja osad ookeani kalad.
Heeringas: Ookeani kala, kes riinvees kasvab väiksena –
kutsutakse räimeks.
Soolsus < 30‰ – soolane vesi
Joogivee soolsus on alla 0,1‰!
Maismaal on soola sisaldus eriti suur kohtadel, kust meri on taandunud ja muld on küllastunud
sooladega. Samuti on suur soolasisaldus kohtades, kus on kõrge põhjavee tase, mis on soolade
rikas.
Eestis soolsus probeelmiks ei ole.
7,4
5-6
2,1
19
4. Informatsioon
Informatsioon on tingimusfaktor juhul, kui see mõjutab organismide elutegevust.
Erinevad infokandjad:
1. FOTORETSEPTSIOON – Võime vastu võtta optilisi signaale erinevates
lainepikkustes. Elektronmagnetkiirguse tajumine.
Silm – kõige võimsam kaheksajalal. Lindudel on silmad väga erineva võimekusega ja
ehitusega
2. MEHHANORETSEPTSIOON – Puutetundlikud retseptorid.
Kaladel on küljejooneelund – sinna on koondunud retseptorid elundiks, mis tunnetab
vee liikumise suunda ja rõhkude vahet.
Putukatel – tundlad
Kassil – vurrud
NB! Nahk pole mehhanoretseptor, sest naha eesmärk on teine.
3. AUDIORETSEPTSIOON – Võime võtta vastu helisignaale, mehhaanilisi võnkumisi.
Bioloogilises maailmas haruldane. Audioretseptsioon on vaid imetajatel, lindudel ja
teatud putukatel. Kehtib reegel: kes teeb häält, see kuuleb häält. Nt: ritsikad
Leidub ka praktiliselt kurte linde – raisakotkad.
Vaid nahkhiired kasutavad kajalokatsiooni. Nad piiksuvad ultraheli alas (inimene ei
kuule) ja keskkonnast tagasi peegeldunud info abil määravad ülimalt täpselt ära saagi
asukoha. Lisaks suudavad nahkhiired vabalt ja kiiresti määrata kõik takistused (tuba täis
niite pole probleem)
4. BARORETSEPTSIOON – Võime tajuda rõhkude vahet.
Oluline vees elavatel organismidel, sest nad üldjuhul liiguvad vees üles - alla. Sellega
kaasneb rõhu vahe ja organismidel on tarvilik siserõhu kohandamine välisrõhuga.
Seetõttu on vaja neil hankida infot välisrõhu kohta.
Kaladel on selleks ujupõis. Ujupõie pinnal paiknevad baroretseptorid.
5. MAGNETRETSEPTSIOON – Võime tajuda magnetvälju.
Magnetretseptsioon on tavaline lülijalgsetel ja lindudel, kellel on magnetvälja jõujoonte
määramiseks vastavad elundid.
Lindudel asub antud elund silma võrkkestas. Sealsed rakud sisaldavad magnetiiti
Fe3O4, mis on kergelt magnetiseeruv aine.
Magnetretseptsioon esineb ka reptiilidel (kilpkonnadel) ja krabidel, kes elavad tõusu-
müüna piiril. Krabid leiavad ülima täpsusega üles mudaväljast oma pesitsuspaiga.
6. ELEKTRORETSEPTSIOON – Võime tajuda elektrivälja.
Kõik organismid tekitavad enda ümber elektrivälja, mida on võimelised tajuma haid ja
raid. Nad kasutavad elektroretseptsiooni mudasse kaevunud saakloomade leidmiseks.
Nad tunnetavad jõudu, mis avaldub kehale elektriväljas – Lorenzi jõud.
Neid organeid nimetatakse Lorenzi ampullideks.
20
KOKKUVÕTE TINGIMUSFAKTORITEST:
1. TEMPERATUUR
2. PH
3. SOOLSUS
4. INFORMATSIOON
4.1 fotoretseptsioon
4.2 mehhanoretseptsioon
4.3 audioretseptsioon
4.4 baroretseptsioon
4.5 magnetretseptsioon
4.6 elektroretseptsioon
21
SÜNERGEETILISED EFEKTID – keskkonnafaktorite koosmõju
• Energia, mis ei avaldu süsteemi üksikkomponentidele
Nt: pannkooki süües tunneme pannkoogi maitset, mitte jahu, muna ja piima maitset.
kahe keskkonnafaktori koosmõju jaguneb:
1. Hädavajalikud ressursid
Nt: vesi ja toitained. Mõlemad on hädavajalikud, kuid veega toitu
(või vastupidi) asendada ei saa.
Puudub keskkonnafaktorite koostoime. Vee ja toidu sünergia on
võrdne nulliga.
2. Asendatavad ressursid
Nt: nisu ja oder. Pole vahet, kas anda tibule nisu või otra – suureks
kasvab ikkagi. Odraga kulub lihtsalt veidi kauem aega.
3. Komplementaarsed ressursid
Ressursi kogukulu on üksi tarvitades suurem, kui koos.
Koostarbimine aitab resurssi kokku hoida.
Nt: hiinlastel – riis. Nad on sunnitud juurde sööma ka loomset toitu,
sest riisis puudub valk. Riisis puudub asendamatu aminohape lüsiin,
mida hiinlased saavad loomsest toidust.
Ameeriklastel – oad. Oad on jällegi valgurikkad. Neis on pea kõiki
asendamatuid aminohappeid (välja arvatud N-sisaldavad tsüsteiin ja
metioniin). Selleks, et kõik vajalikud aminohapped ubadest kätte
saada, on neid vaja süüa kaks korda rohkem, kui energeetiliselt vajalik.
Lahendus: Sojauba transporditakse siiani Ameerikast Hiinasse suurtes kogustes ning Ameerika
töölistele antakse riisi.
Koos tarbides on ressursside kogukulu väiksem.
4. Antagonistlikud ressursid
Koos tarbides ressursside kulu suureneb. See on seotud
herbivooridega ja tingitud faktist, et taimed toodavad toksiine, et neid
ära ei söödaks.
Troopikas on täheldatud, et taimed teevad toksiinide tootmisel
tahtmatultevolutsiooniliselt koostööd. Nt: Akaatsiate mürgid üksi on
loomadele kahjutud, kui aga tarbida eCrinevaid akaatsiaid korraga,
on toksiinid surmavad. Nii vajab loom veel lisatoitu, et käivitada
mürkidest vabanemiseks biokeemilisi reaktsioone.
22
TOLERANTSIKÕVER JA ÖKOLOOGILINE NIŠŠ
Kõiki organisme iseloomustavad kohastumised – fülogeneetiline adaptsioon.
Kohanemine on ontoloogiline adaptsioon, ilma geneetilise muutuseta.
Inimesel pole kohastumust hairünnaku puhul, kuid intuitiivselt teame, kuidas käituda nt
karurünnaku puhul.
Tolerantsikõver – näitab, kuidas on organismi heaolu seotud ökoloogilise faktoriga.
Tolerantsikõver = gaussi kõver.
Faktor – Olulise ökoloogilise faktori tase
(mulla niiskus, pH, jne)
Optimum – optimaalne faktori tase, mis
vastab organismi kohastumisele.
C – maksimaalne elutegevuse intensiivsus,
milleks vaadeldav liik on võimeline.
Tolerants – vahemaa miinimum ja
maksimum faktori väärtuse vahel, kus liik
on
võimeline
elama.
Ökoloogiline
amplituud – näitab, kui laias faktorite
väärtuste vahemikus suudab taim toime
tulla.
Organismid jagunevad:
Stenotoop – spetsialistid, kitsa ökoloogilise amplituudiga
Eurotoop – elavad kõikjal kus vähegi võimalik, laua ökoloogilise amplituudiga.
Nt: kuusk ja mänd
Kui kuusk ja mänd kasvavad samal alal, surub kuusk männi
alla.
Ökoloogiline nišš = kohastumuste kompleks
• Liigi omadus, mis määrab, mis rolli liik evolutsioonis täidab.
Ökoloogiline nišš on see osa n-mõõtmelisest faktorruumist, mille telgedeks on olulised
ökoloogilised faktorid kus liik suudab edukalt eksisteerida, ehk tingimused, millele liik on
kohastunud.
n = faktorruumi mõõtmelisus
n = 1. Ökoloogilist amplituudi saab vaadata kui ühemõõtmelist nišši.
23
n = 2. nt: kuuse ja männi 2-mõõtmeline nišš.
Märgitakse ringina, sest äärealadel on mulla niiskus ebasoodne ja
pH ebasoodne.
Ring = Hilberti ruum. Hilberti ruumis kehtivad Eukleidese
seadused.
n = 3
Kuna Hilberti ruumis kehtib Eukleidese matemaatika, siis distantsi
saab alati leida, olenemata n-ist.
d = (F12 + ....... + Fn2)0,5
Kuigi me teame, kus mingi taim võiks elada, siis see ei tähenda, et ta seal elab.
Konkuretns/sööjad/parasiidid ei võimalda liikidel igal pool elada, kus nad võiksid. Taim ei ole
igal pool konkurentsivõimeline ja tihti sunnitakse ta taanduma palju väiksemalt alale.
Siit tuleneb:
1. Fundamentaalne ehk autoökoloogiline nišš – ala, kus taim võiks elada
2. Realiseerunud ehk sünökoloogiline nišš – kus taim tegelikult elab. alati väisem kui
potentsiaalne nišš.
Nt: taim, kes looduses elab ainult liival, kasvab ka rammusas peenras. Looduses aetakse ta
rammusest mullast ära.
Nt: Mändi leidub oma fundamentaalse nišši keskel vähe, sest kuusk ajab ta sealt ära.
NB! Nišš on liigi omadus! Keskkonnas nišše pole, keskkonnas on faktorid!
24
ÖKOLOOGIA SEADUSED
1. Liebigi seadus: Organismide kasvu ja paljunemist limiteerib see faktor, mis on liigi
optimumist suhteliselt kõige kaugem.
„Limiteeriv ressurss“ – termini tõi kasutusele Justus von Liebig.
Alati leiab üles kõige limiteerivama ressursi, mis limiteerib toodangu.
Nt: Päevalillel on kaaliumi puudus limiteeriv ressutss – Nt naatriumi juurde andes
kaaliumipuudust ei paranda.
2. J.Shelfordi seadus – on täiustatud Liebigi seadus.
Tee miinimumini on erinev.
WTF TÕLGI
25
FENOTÜÜBILINE PLASTILISUS
Fenotüüp – genotüübi realisatsioon. Organismi vaadeldavate/mõõdetavate tunnuste kogu, mis
kujuneb välja ontogeneesi (organismi individuaalse arengu) käigus. Eelkõige mõjutab
fenotüüpi genotüüp, aga ka konkreetne keskkond, kus organism elab.
σF2 - Kogu fenotüübiline varieeruvus (mida kohtame nt populatsioonis). (dispersion)
σF – standardhälve
Kogu fenotüübilist varieeruvust mõjutab:
• Genotüüp σG2 - Peamõju
• Keskkonna tekitatud mõju σK2 - Peamõju ! Plastilisus väljendub
• Keskkonna ja genotüübi koosmõju σK*G2 - Koosmõju ! Plastilisus väljendub
• Viga, juhuslikkuse component σE2 - Juhuslik varieeruvus
σF2 = σG2 + σK2 + σK*G2 + σE2
-------------------- ----------- ---------
PEAMÕJU KOOSMÕJU JUHUSLIKKUS
Plastilisuse väljendumine: Adaptiivne plastilisus
nt: Harilik mänd.
1) Lageraiesmiku männid.
Kogu energia on männid kulutanud pikkuskasvuks, sest muidu jääksid nad
teiste varju, pimedusse ja sureks.
Aktiivne plastilisus!
See on mändide püüe parandada oma fitnessi, ehk kohasust.
Kohasus – lühiajaline paljunemisedukuse mõõt.
2) Looduskaitse all olev mänd.
Ta on kasvanud üksi, ilma konkurentsita PARi pärast. Selleks,
et rohkem PARi saada, on mõttekam lateraalselt levida.
Plastilisuse väljendumine: Passiivne plastilisus
Fenotüübi muutused ei pea alati olema seotus kohasuse muutusega.
Nt rabamänd on väike, see on paratamatu reaktsioon, sest toitaineid pole
Plastiline organism muutub ontogeneesi käigus, aga keskkonna
tagasimuutumisel, ei muutu organism tagasi.
26
Fenotüübiline elastsus – keskkonnatingimuste tagasi muutumisel, muutub organism tagasi.
nt: jänese karv talvel ja suvel
nt: päevitades muutub meie nahk tumedamaks, kui päike kaob ära – nahk tagasi valgeks.
:) Meie oleme valged, et rohkem D-vitamiini kätte saada.
Kuidas plastilisust ära tunda?
reaktsiooni norm – eeskiri, kuidas keskkond fenotüüpi mõjutab.
σG2 + σK2 Peamõju – keskmine keskkonna mõju
σG2 > 0: On olemas 2 erinevat (geneetilist) fenotüüpi (punane,
sinine) – on olemas geneetiline varieeruvus
σK2 > 0: On olemas keskkonna mõju, mis muudab genotüüpi
ühesuguse intensiivsusega. – Ühesugune intensiivsus = ühesugune
tõusunurk.
σK*G2 = 0: Koosmõju pole, sest keskkond ei mõjuta fenotüüpi
erineval moel.
σG2 > 0: On olemas 2 erinevat (geneetilist) fenotüüpi (punane,
sinine) – on olemas geneetiline varieeruvus
σK2 > 0: On olemas keskkonna mõju, mis muudab genotüüpi
ühesuguse intensiivsusega. – Ühesugune intensiivsus = ühesugune
tõusunurk.
NB!
σK*G2 > 0: siin on olemas ka koosmõju, keskkond mõjub
ühele genotüübile intensiivsemalt kui teisele.
Plastilisus puudub: tegu on fikseeritud tunnusega. nt – ninade arv.
Evolutsioonis puudub mõte rohkem kui üht genotüüpi üleval hoida.
σG2 + σK2 Peamõju = 0
27
DEMÖKOLOOGIA – populatsiooni ökoloogia
Demökoloogia eesmärk on selgitada, mis asjaoludel populatsioonid tekivad, püsivad, kasvavad
ja hääbuvad. Need on 4 faasi, mille iga populatsioon läbi teeb.
Bioloogilise liigi keskmine eluiga on 106 aastat.
N – Populatsiooni tihedus.
Konstantsel pindalal on populatsiooni tihedus ekvivalentne liikmete arvuga.
Tihedust tunnetab iga isend iga oma keharakuga.
Millest populatsiooni tihedus (N) sõltub?
Võrrand on parameetriteta.
Parameeter – mingi üldistav suurus. Võrrandites muutujate kordajad mis näitavad, mis
intensiivsusega mingi jõud võrrandit mõjutab.
Npraegu
= Nenne + sünnid – surmad + immigratsioon – emigratsioon
Nenne – Eelmine ajavahemik, mis pani baasi praegusele olukorrale.
Nt: Euroopa naaritsa populatsiooni suurus Hiiumaal. N praegu = N enne + Sünnid – surmad
(ei ole rännet, sest naaritsad asuvad saarel)
Üksiku populatsiooni piiratud kasvuvõrrand
Veidi kontekstist välja tiritud, arvestab vaid populatsioonisiseseid protsesse.
Üksiku populatsiooni kasvuvõrrand näitab, kui palju populatsiooni tihedus ajaühikus muutub.
dN/dt – populatsiooni muutumise kiirus.
dN/dt > 0 – populatsioon kasvab
dN/dt < 0 – populatsioon kahaneb
dN/dt = 0 – populatsiooni tihedus on ajas stabiilne
dN/dt = B(birth) – D(death)
B = bN
D = dN
b,d – sõltuvad parameetrid.
dN/dt = bN – dN
dN/dt = N (b – d)
b - d = r – vastava vaadeldava liigi biootiline potentsiaal = erikasvukiirus. Iseloomustab,
millise kiirusega tihedus mõjutab kiirust. Suur r = kiire paljunemine!
dN/dt = Nr – populatsiooni piiramatu kasvu võrrand. MALTHUSE VÕRRAND.
Populatsiooni eksponentsiaalne kasv.
28
T.H.Malthus oli teadlane kes selle võrrandi põhjal püstitas hüpoteesi, et sõjad on paratamatud
ja hädavajalikud inimeste arvu vähendamiseks.
Populatsiooni kasvu pidurdav jõud – keskkonna takistus.
Kui populatsiooni arvukus muutub liiga suureks, on paljunemine prsitud.
Nt: Haiguste levik
võistlus ressursside pärast
Kuidas on populatsiooni kasvu pidurdav jõud seotud tihedusega?
R = a N2 Takistus on tiheduse ruut läbi mingi parameetri a.
Maismaal on üldiselt resurss kahemõõtmeline (põllumaad mõõdetakse
ruutmeetrites, mitte kuupmeetrites)
Tihedus ja takistus on seotud paraboolselt populatsiooni tihedusega.
K – keskkonna kandevõime. K on suuresti liigi omadus. Keskkonna kandevõime näitab, kui
palju suudab keskkond vaadeldavat liiki taluda – Maksimaalne vastava populatsiooni tihedus
vaadeldavas keskkonnas. (Inimeste K on väga suur!)
K = r / a
r – Paljunemise mõju populatsiooni kasvule. Suuresti liigi omadus!
a – millest suurem isendite arv populatsiooni kasvu takistab, eeldusel, et liigisisene konkurents
on põhiline.
Konkurents on (- -) interaktsioon. Vastasmõju, mis mõlemale osapoolele mõjub
negatiivselt. Konkurents on kahepoolselt kahjulik vastasmõju.
K = r / a => a = r / K
dN/dt = rN -aN2 = rN – (r/K)*N2
dN/dt = rN (1-N/K)
----- ------------
osa, mis vaid kasvab pidurdav jõud
dN/dt = rN ( 1 – N / K) – populatsiooni piiratud kasvu võrrand. (Logistilise kasvu
võrrand/ sigmoidse kasvu võrrand)
Malthuse võrrand vs populatsiooni piiratud kasvu võrrandi graafikud
Carrying capacity = asümptood K
N
R
29
Võrrandil on revolutsiooniline sisu:
§
2 muutujat: N ja t (tihedus ja aeg)
§
2 parameetrit r ja K – (liigi biootiline potentsiaal ja keskkonna kandevõime. - eelkõige
LIIGI omadused.
Looduses eristatakse:
r-stateegid: Panus suurele järglaste arvule. Kiire võime resursse hõivata.
k-stateegid: Ressursside tarbimine väga efektiivselt, suudavad säilitada efektiivselt suurt
populatsiooni tihedust.
30
Üksiku populatsiooni piiratud kasvu võrrand
Populatsiooni ajalise dünaamika vaatlemisel on keskse tähtsusega populatsiooni liikmete arvu
ehk populatsioonitiheduse (N) muutus ajas (t) ja samuti muutumise suund, mida väljendab
populatsiooni kasvukiirus ΔN/Δt, diferentsiaalkujul dN/dt. Kui populatsioon ajas suureneb,
on kiirus positiivne, vähenedes negatiivne. Nullkasv (dN/dt = 0) tähendab populatsiooni
liikmete arvu püsimist samal tasemel. Jättes lihtsuse mõttes tähelepanuta immigratsiooni ja
emigratsiooni, saab kasvukiirust väljendada ajaühikus sündivate ja ajaühikus surevate isendite
arvu vahega (vastavalt B ja D)
dN/dt = B – D.
Sündide ja surmade koguarv sõltub seejuures mõistagi populatsioonitihedusest – mida rohkem
isendeid, seda rohkem sünde ja seda rohkem surmasid toimub ajaühikus. Olenevalt liigi
paljunemiskiirusest ja suremusest tuleb siin mängu võtta ka liigiomased sünni
- ja
surmakoefitsiendid ehk
-parameetrid (vastavalt b ja d):
dN/dt = bN – dN.
Tuues parameetrid sulgude ette ja tähistades nende vahet uue liigiomase koefitsiendiga
r = (b – d),
saame: dN/dt = rN,
kus r on erikasvukiirus ehk sisemine kasvukiirus ehk biootiline potentsiaal, mis näitab liigi
võimet ajaühikus ühe populatsiooniliikme kohta järglasi toota. Parameeter r on alati positiivne
arv, sest igal bioloogilisel liigil on potentsiaali rohkem lapsi teha kui isendeid vanaduse tõttu
loomulikult sureb. Võrrandit (7) nimetatakse populatsiooni piiramatu kasvu võrrandiks ehk
populatsiooni eksponentsiaalse kasvu võrrandiks ehk Malthuse võrrandiks. Thomas
Robert Malthus (1766
-1834) oli Inglise demograaf ja majandusteadlane, kes aastal 1798
avaldas kuulsa essee, kus vältis, et vaesus ja sõjad on paratamatud, kuna elanikkond kasvab
geomeetrilises, elatusvahendite hulk aga aritmeetilises progressioonis. Seda väidet
illustreeribki piiramatu kasvu võrrand, kust nähtub, et populatsiooni kasv on autokatalüütiline
ehk positiivselt tagasisidestatud protsess – „mida rohkem, seda rohkem; mida vähem, seda
vähem“ (negatiivne tagasiside tähendab regulatsiooni: „mida rohkem, seda vähem; mida
vähem, seda rohkem“).
Ilmselgelt ei saa üksiku populatsiooni kasv tegelikkuses olla piiramatu – ressursse on
ökosüsteemis alati piiratud hulgal ja mingi populatsioonitiheduse juures hakkab ressursipuudus
kasvu piirama, s.t. keskkond hakkab avaldama populatsiooni kasvu pidurdavat mõju, i.e. tekib
nn keskkonnatakistus (R). Järgnevalt olekski vaja sedastada, mil viisil võiks seda pidurdavat
mõju matemaatiliselt formuleerida. On põhjust eeldada, et keskkonnatakistus on seotud
populatsioonitiheduse ruuduga. Nimelt vaadeldakse eelnevates võrrandites populatsiooni
31
liikmete arvu mingi ressursse kandva pindalaühiku kohta. Pind on kahemõõtmeline ruum,
organismid on aga kolmemõõtmelised. Seega:
R = aN2,
Kus a on liigi
- ja keskkonnaomane parameeter. Lisades Malthuse võrrandi paremale poolele
miinusmärgiga keskkonnatakistuse, saame:
dN/dt = rN
- aN2.
Defineerides läbi parameetrite r ja a uue liigiomase parameetri K = r/a, ja nimetades selle
keskkonna
kandevõimeks, ning tuues korrutise rN sulgude ette saame:
dN/dt = rN(1 – N/K) = rN((K – N)/K).
Keskkonna kandevõime K näitab, kui suurt vaadeldava liigi populatsiooni on keskkond
võimeline kandma, ehk siis majutama ja ära toitma. Saadud võrrandit nimetatakse
populatsiooni piiratud kasvu võrrandiks ehk logistilise kasvu võrrandiks. Näeme, et
populatsiooni kasvukiirus on positiivne senikaua, kui N < K. Tingimusel N = K on populatsioon
nullkasvus (N on ajas stabiilne), kui N > K on kasv negatiivne, st populatsiooni isendite arv on
languses. Matemaatilises mõttes on keskkonna kandevõime seega populatsiooni maksimaalne
võimalik stabiilne tihedus ehk populatsioonitiheduse asümptood. Viimast saab märkida
populatsioonitiheduse teljele graafikul, kus piiratud kasvu võrrand on integraalkujul esitatud.
Piiratud kasvu võrrandis on seega kaks muutujat (N, t) ja kaks liigiomast parameetrit (r, K).
Viimased on olulised liigi põhilisi kohastumusi väljendavat suurused. Kui r on kõrge, siis liik
on kiire paljuneja, kui K on kõrge, siis liik kasutab ressursse efektiivselt ja ökosüsteemis on
korraga võimelised elatuma ning paljunema suur hulk isendeid. K kujuneb evolutsioonis välja
kui sündivuse ja suremuse tasakaal teatava populatsioonitiheduse juures. Mõlemad võivad
sõltuda N
-st ja mõlemad võivad olla ka N-st sõltumatud.
32
LIIGISISENE KONKURENTS – kahepoolselt kahjulik biootiline vastasseis.
- konkurents lõplikule ressursile
Suurimad konkurendid on üldiselt liigisiseselt.
Konkurents jaguneb:
1. Sümmeetriline konkurents – kõik kaotavad võrdselt
2. Asümmeetriline konkurents – Joonistuvad välja võitjad/kaotajad nt: valguskonkurents
Varjestatav vs mittevarjestav konkurents
• Valguskonkurents: Suur varieeruvus, Pisike algedu viib suure ülekaaluni. PAR on
varjestatav ressurss.
• Juurte konkurents: varieeruvus on väiksem, kõik kaotavad võrdselt/proportsionaalselt.
Mullaresursid ei ole varjestatavad.
J.Weineri liigisisese konkurentsi eksperiment.
Valge – ühe taime keskmine mass
Punane – Taimede massi variatsioonikoefitsent.
33
KONSTANTSE SAAGI SEADUS
= ISEHÕRENEMISE SEADUSE ERIJUHT
= -3/2 ASTME SEADUS
Konstantse saagi seadus käib peamiselt põllukultuuride kohta, kus saaki korjatakse kogu
biomassiga (söödakultuurid, nt ristik).
Kogu saak ei sõltu istutamistihedusest.
Konstantse saagi seaduse kontroll ristiku ja luste (kõrreline) liigiga.
x-telg: külvitihedus
y-telg: saagiks saadud biomass pindalaühiku kohta
Saagikus paraneb kui on alla 2500 seemne, kuid sealt edasi on lisakülv seemnete raiskamine.
Saagikus paraneb ka viljakama mullaga.
W – ühe taime keskmine mass
N – taimede arv pindalal
kogu taimede mass, WN = const (c)
W=cN-1 => logW = logC – 1logN
hüperbool logaritmidega taandub lineaarseks:
Konstantse saagi seadus on üldisema seaduse – isehõrenemisseaduse – erijuht.
Kohort – ühevanune osa populatsioonist.
nt: raiesmikule tihedalt istutatud kuused. Seal on liigisisene konkurents suur, kus enamus sureb.
Edukamad haaravad resursi – ruumi. Tekib konkurentsi poolt tekitatud suremus.
Järeldus - metsakihtide arv peab olema lõplik suurus! Mets koosneb lehekihtidest ja kuna ka
kõige alumina kiht peab saama valgust – peab olema lõplik suurus. Kui kihte oleks lõputult –
alumised kihid valgust ei saaks.
34
Isehõrenemise seadus ja selle eeldused.
1. eeldus
(specific leaf area) SLA = lehepind / pindalaühik = L
L = const.
1. eeldus
L = λN = const
λ – 1 isendi keskmine lehepind
N – polulatsiooni tihedus
2. eeldus: 1 taime lehtede kogupindala λ on seotud taime diameetri ruuduga (läbi mingi
koefitsiendi, keskkonna parameetri a)
Taimede projektsiooni diameter annab taime pindala
2.eeldus
λ = aD2
3. eeldus. saab esitada 2 erineval kujul, sõltuvalt sellest, kuidas kujutame täiskasvanud
populatsiooni.
1) 1 taime massi ja diameetri sõltuvuse olemus (3D)
W – taime keskmine mass
3.eeldus
W = bD3
2) taimel puudub kõrgusmõõde (2D)
3.eeldus
W = bD2
Meid huvitab populatsiooni tiheduse (N) ja taime kesmise massi (W) vaheline seos.
W = bD3 puhul:
W = cN-3/2
(W = cN-3/2 => logW = logc – 1,5logN)
W = bD2 puhul:
W = cN-1
(W = cN-1 => logW = logc – logN)
See on isehõrenemise seaduse erijuht, kui taimed on pigem 2D
35
Isehõrenemise seadus oma algsel kujul vs konstantse saagi seadus graafik
Isehõrenemise seaduse, ehk -3/2 astne seaduse illustratsioon
x-telg: külvitihedus (log)
y-telg: isendi keskmine biomass (log)
KASVU LAGI
Taime massi kasvades, tihedus väheneb.
Taimed saavad kasvada vabalt (kasvu laeni), kuni
kasvavad piisavalt suureks (et oksad hakkaksid koku
puutuma), et tekiks valguskonkurents.
Mida hõredamalt on istutatud taimed, seda kõrgemal
on kasvu lagi (seda kauem saavad kasvada ilma
konkurentsita).
Väike algedu võib tähendada suurt lõppedu.
Osadel taimedel piisavalt suureks saades -3/2 seadus murdub konstantse saagi seaduseks. See
juhtub, kui taimed enam suuremaks ei kasva ja taimed hakkavad kõrgusesse pürgimise asemel
laiutama – D3 asendub D2.
D2 taimede kasv
!Just valguskonkurents tekitab seose -3/2. Kui valguskonkurents puudub, kehtib seos -1!
36
Konstantse saagi seadust on taimekasvatusega, eriti aga söödataimede kultiveerimisega
tegelevad põllupidajad intuitiivselt ja empiiriliselt teadnud juba ammusest ajast
-- seemet ei
maksa väga palju raisata, optimaalsest tihedam külv või istutamine üldjuhul saaki suurenda.
Teisisõnu, olenemata külvitihedusest tuleb saak ikka samasugune. Seda mõistagi ainult siis kui
taimed jagavad ühist ressurssi, mitte ei kasva eraldi pottides või üksteisest kaugel.
Konstantse saagi seaduse saab üles kirjutada järgmiselt, defineerides saagi kui ühe täiskasvanud
taime keskmise massi (w) korrutatuna isendite arvuga pindalaühiku kohta (N):
wN = const → w = const/N;
tähistades konstandi lühemalt c
-ga saame:
w = cN-1.
Pärast mõlema võrrandipoole logaritmimist saame konstantse saagi seadust tähistava lineaarse
võrrandi:
logw = logc
- logN.
Näeme, et tegemist on langeva sirge võrrandiga, mille tõus on
-1 ja vabaliige võrdne log c-ga.
Paraku on ökoloogia praktika näidanud, et looduslikes populatsioonides, kus kohordi
sirgumisega kaasneb märkimisväärne suremus, st toimub kohordi isehõrenemine, konstantse
saagi seadus enamasti ei kehti, vaid log w ja log N seos on järsema langusega kui
-1 (s.t. sirge
tõus <
-1). Täpsemal hindamisel on selgunud, et sellisel juhul on sirge tõus enamasti võrdne -
3/2
-ga.
Püüame konstantse saagi seadust ja isehõrenemise seadust teoreetiliselt tuletada, toetudes
mõnele lihtsustavale eeldusele. Selleks toome kõigepealt mõistena sisse lehepinna indeksi (L,
tähistatakse tihti ka lühendiga LAI, ingliskeelsest terminist Leaf Area Index) mõiste.
Meenutuseks, L on dimensioonitu suurus, mis näitab kui palju on taimepopulatsioonis kokku
lehepindala maapinna ühiku kohta (näiteks lehepinna ruutmeetreid ühe maapinna ruutmeetri
kohta).
Esimene lihtsustav eeldus väidab, et L ei kasva lõputult koos taimede massi suurenemisega vaid
jääb mingist kohordi arengujärgust alates konstantseks. Seda lihtsal põhjusel, et lehestik ei saa
koosneda lõputa paljudest PAR
-i püüdvatest kihtidest, alumistele lehtedele sellisel juhul
valgusressurssi ei jätkuks. Seega:
L = λN = const;
kus λ tähistab ühe täiskasvanud taime keskmist lehepindala.
Teine eeldus väidab, et ühe taime lehepindala (juhul kui L
-i maksimumväärtus on saavutatud
ja lehestiku kihtide arv lõplik ning kohordi jaoks konstantne) on seotud taime poolt hõlvatud
ruumiosa diameetriga läbi ruutseose:
37
λ = aD2,
kus D on taime poolt hõlvatud pindala diameeter ja a on liigiomane koefitsient (konstant).
Eeldus on igati mõistlik
-- kui lehestiku kihtide arv ja seega L, on konstantne, suureneb lehepind
taimel ainult diameetri kasvu arvelt, pindala on aga teatavasti diameetri ruudu funktsioon.
Kolmas eeldus väidab, et ühe keskmise taime mass on taime diameetri kuubi funktsioon:
w = bD3,
kus b on liigiomane konstant. Selline eeldus peaks kehtima juhul kui taim kasvab laienemisega
paralleelselt ka kõrgusesse ja täidab hõivatud kolmemõõtmelise ruumi enam
-vähem ühtlaselt
biomassiga, mitte ei kasuta väetimate naabrite allajäämist konkurentsis ainult laiutamiseks.
Teisisõnu, kui taime saab võrrelda silindri või kuubikuga, mitte näiteks taldrikuga. Kuubiku ja
sellesse mahtuva silindri puhul on mass ju tõepoolest diameetri kuubi funktsioon.
Aga, teatud juhtudel, eriti põllumajanduses ette tulevail, on taimed aretatud selliseks, et nad
väga kõrgusse ei pürgigi ja pigem püüavad kasvada laienemise arvelt. Sellisel juhul on mõistlik
kolmandat eeldust muuta (20) ja eeldada ruutsõltuvust:
w = bD2.
Kui nüüd ülalpool formuleeritud eeldusi kasutades hakata tuletama w Ja N seost, saades
asendamise teel lahti nii D
-st kui λ-st, selgub, et eelduse w = bD3 rakendamine viib meid
võrrandini:
w = b(L/a)2/2 N-3/2.
Näeme, et võrrandi parema poole avaldis võrdusmärgi ja N
-i vahel koosneb ainult konstantide
kombinatsioonist (b, L, a), seetõttu võib seda osa võrrandis tähistada lihtsalt c
-ga:
w=cN-3/2 →logw=logc
-1,5logN
See ongi isehõrenemise ehk
-3/2 astme seaduse matemaatiline väljendus.
Kui aga kasutada kolmanda eeldusena seost w = bD2 saame pärast asendamisi:
w = b(L/a)2/2 N-2/2.
Tähistades ka nüüd võrrandi paremal poolel leiduva konstantide kombinatsiooni üheainsa
sümboliga (c), saame:
w=cN-1 →logw=logc–logN.
Seda võrrandit tunneme kui konstantse saagi seaduse matemaatilist kuju.
Nagu eelpool näha, sõltub populatsioonitiheduse ja ühe taime massi vaheline seos sellest,
milline on taimede kasvustrateegia ja kasvuviis. Mida "kahemõõtmelisem" on taim seda
paremini kehtib konstantse saagi reegel, mida "kolmemõõtmelisem" on taim, seda paremini
kehtib
-3/2 astme ehk isehõrenemise reegel.
38
Mitmed uurimused on näidanud, et kohordi arengu küpses faasis isehõrenemise seadus üldjuhul
"murdub" konstantse saagi seaduseks
-- täiskasvanud taimed kasvavad lõpuks paljude naabrite
surma tõttu peamiselt laiusse ja w edasine muutus on siis ennustatav pigem läbi taime diameetri
ruudu, mitte läbi diameetri kuubi.
39
POPULATSIOONI DEMOGRAAFILISED PARAMEETRID
- kirjeldavad liigi/keskkonna-omaseid protsesse
- peegeldavad protsesse, mille toimimisel populatsioonid sünnivad, püsivad, kasvavad ja
surenav
- populatsiooni dünaamikat kirjeldavad üldistused
Populatsiooni uurimisele saab läheneda 2 moodi:
1) Kohordi meetod = Dünaamiline meetod
Jälgitakse 1 põlvkonna elu sünnist surmani.
Seda on agar aske uurida, sest paljud ligiid elavad meist kauem. nt: polaarhai elab 400
aastaseks.
2) Staatiline meetod = Rahvastiku püramiidi meetod
Järeldusi tehakse populatsiooni demograafilise vanuselise struktuuri kohta.
nt: kui palju lapsi läheb üks aasta kooli – järeldus: 7 aastat tagasi tehti palju lapsi.
1) KOHORDI MEETOD
-eelistatavam
Meil on teada liigi arengustaadiumid: muna, vastne (4), Valmik
44000 muna –> 1300 valmikut. Vaid 3% munadest saab suguküpseks.
R0 – PUHAS KASVUKIIRUS. keskmine järglaste arv, mida 1 populatsiooni liige elu jooksul
toodab
R0 = lxmx1+…..+ lxmxn
R0 = 0+0+0+0+0+0,51 = 0,51
VILJAKUS
ELUMUS.
KOHORDI
TÕENÄOSUS
SAAVUTADA
MINGI VANUS
R0 < 1: popul. kahaneb
R0 = 1: stabiilne
R0 > 1: popul. kasvab
40
parameeter T – generatsiooni aeg, eluiga.
- Aeg, mis kulub 1 generatsiooni möödumiseks.
- millise vahega 1 kohort järgneb teisele
dN/dt = rN – (1 – N/K)
r – biootiline potentsiaal/kasvukiirus. Kajastab liigi võimet järglasi toota ajaühikus.
r = ln R0 / t
NB! Elumus (lx) ja puhas kasvukiirus(R0) on hästi olulised! :)
vx – paljunemise väärtus. Näitab tulevase põlvkonna ootust mingis vanuses.
Oleme x vanad:
Näitab kui mitut last on selle kohordi liikmel veel oodata
Näitab, millise panuse kasvu elu jooksul see liige veel annab
VALEM
Sama vanusega populatsiooni liikmed ei ole sama väärtuslikud. Just 20-aastased naised on
populatsiooni taastootmise kohapealt kõige väärtuslikumad.
41
2) STAATILINE MEETOD.
ELUMUSE KÕVERAD EHK LIIGI KÕVERAD
3 tüüpi:
1. Alguses sureb vähe, suremus koondunud
kõrgesse ikka.
Populatsioonis suur vanem pool.
nt: inimene
Suur K! – enamvähem kõik kasvatatakse
suureks.
2. Suremus
on
vanusest
sõltumatu.
Surmarisk on alati sama sugune.
Suremine juhusliku õnnetuse tagajärjel.
3. Suremus koondunud noorukiikka.
Kui juba suureks kasvatud, on suremus väiksem.
nt: lõhe – teeb lapsi tohutul hulgal, suureks saavad vähesed.
suur r!
2) Mingi aja jooksul sureb alati mingi kindel protsent
olemasolevatest. Suremus on vanusest sõltumatu.
Demograafiline püramiid
Prantsusmaa
Näitab ühe sünniaastaga isendite hulka. Ülespildistud mingil ajahetkel populatsiooni kohta.
Augud:
1926 – I MS
1945 – II MS
1980 – seksuaalrevolutsioon. Rasestumisvastased vahendid.
Näha on ka sõjajärgsed beebibuumid.
Analüüsides demgraafilisi püramiide saab välja
lugeda populatsiooni tuleviku.
42
LEVIK JA LEVIMINE
- kahel terminil tuleb selgelt vahet teha
Levik – Isendite paiknemine ruumis üksteise suhtes
Levimine – Protsess, isendite ümberpaiknemine ruumis
1) LEVIK
Areaal. Igal liigil on oma areaal, mis määrab liigi asukoha biosfääris.
Ruumilise leviku tüübid:
a) juhuslik. Isendi asukoht ei sõltu teise isendi asukohast.
umbrohi
vetikad
võilill
b) Regulaarne. Vahemaa isendite vahel on maksimaalne.
Liigid, kes kaitsevad oma territooriumi – nt metsalinnud
c) Agregeeritud. Isendid paiknevad rühmadena. Rühmade vahe on kas reguleeritud või
juhuslik.
Põhjused:
1. Paljunemisviis:
Klonaalselt paljunevate liikide järglased on lähedal.
Sessiilne – substraadile kinnitunud organismid elavad ühes kohas (taimed)
2. Sotsiaalne koondumine: Sotsiaalsed rühmad
Ühiselulised puhukad
Inimesed külas
šimpansite kari, kus on sotsiaalne hierarhia ja tööjaotus
3. Isekus.
nt: sebradel saab tugevaim rühma keskele. Lapsed ja emad on välisringis. Lapsi saab
juurde teha, 1 lapse eest elu anda pole mõistlik. Iga sebra tahab et tema ja lõvi vahele
jääks teine sebra.
43
2) LEVIMINE
Levimine on hädavajalik omadus kõigile organismidele.
Kõik organismid seda ka suuremal/vähemal määral teevad.
Levimiskaugus = kohastumus (või ka selle puudumine).
Levimine:
1. PASSIIVNE
Kõik juhud, kus organism tervikuna üheski staadiumis keskkonnas ei liigu. Levimisteenust
osutavad kas füüsikalised jõud või teised bioloogilised ligiid.
Levimisvektor – Jõud, mis levimist võimaldab.
Passiivse levimise levimisvektorid on:
a) biootilised – Koostöö loomadega (loomad söövad taime seemed koos marjaga ja jätavad
uude kohta koos väetisega), Putukad kannavad õietolmu
b) Tuul (võilill), Õietolm (spermid), Käbitolm (männil), Vesilevijad (taimede seemed
suudavad ujuta vee peal)
Üherakulised eosed on levimise maailmameistrid (sõnajalaeosed, orhideede seemed, mis on
massi poolest võrreldavad eosega)
Priimula nt aga loodab ainult lülijalgsete eale, keda ta seemed huvitavad ja kannavad piisavalt
hästi laiali.
2. AKTIIVNE
Organism suudab ise ruumis levida ja partnerit otsida.
Nt: kaheksajala isased kasvatavad sugujala, mis suudab levida kiiresti ja läheb kaaslast otsima.
Kaugele levik on evolutsiooniliselt oluline. Levimine on hädavajalik, sest evolutsiooniliselt
pole hea, kui järglasi saadakse lähisugulastega.
Geneetiline degratsioon – geenilookused muutuvad homosügootseks. Tekib inbriidingu
depression.
Kehvem immuunsus
Kehvem fenotüüp
Halb on saada lapsi ka geneetiliselt väga kaugete liigikaaslastega. Tekib vastupidine efekt –
outbriiding.
Samad alleelid peaaegu et puuduvad.
Kehvem immuunsus
Vana-Egiptuse valitsejate keskmine eluiga oli 15 aastat. See on inbriidingu depressiooni
tulemus.
Lindudel (nt rasvatihastel) tagab kodupesast 1km kaugusele rändamine inbriidingu ja
outbriidingu puudumise. Kusjuures kaugemale levivad rohkem emased (Y kromosoom levib
aeglasemalt kui X)
44
MIGRATSIOON – Levimise erivorm
Suure hulga isendite suunatud liikumine ühest kohast teise.
Migratsiooni tüübid:
1. Korduvkasutusega pilet
Tavaline interopaarsetele liikidele – korraga saadakse vähe lapsi, mitu paljunemispingutust.
• Rändlinnud
• Suurte mereimetajate ränded (rändkilpkonn, vaal)
Põhjustab: aastaaegade vaheldumine
2. Edasi-tagasi pilet
Migreerutakse toitumispaika ja üks kord paljunemispaika. Tavaline semelpaarsetele liikidele
– elu jooksul toimub 1 paljunemispingutus.
nt: sääsed. Sünnivad/kasvavad lombis ja siis lähevad verd imema. Naasevad paljunemispaika,
munevad ja surevad
nt: vaikse ookeani lõhelised. Sünnivad jões, toituvad ookeanis ja paljunevad jões ning surevad.
3. 1-otsa pilet
Väga haruldane. Migreerutakse 1 kord, tagasi migreerivad lapsed.
NB! Täheldatud ainult liblikatel (admiral, monarh).
Nt: Monarh Põhja-Ameerika Lousiana kandist milonitega rändavad Niššigani kanti, munevad
ja surevad. Järglased rändavad tagasi.
Eelis: Ükski loom ei spetsialiseeru sellise liigi toitumisele.
Taimede puhul:
Semelpaarne = monokarpne
Interopaarne = polükarpne
Demökoloogia lõpp
45
KOOSLUSÖKOLOOGIA – SÜNÖKOLOOGIA
Biootiliste interaktsioonide vastastikmõju.
Eeldus: Omavahel suhtlevad liigis omavad vastastikmõju
Interaktsioonid:
1. Neutraalne (0,0) – Ei avaldata olulist mõju
2. Konkurents (-,-) – Kahepoolselt kahjulik.
a. Liigisisene
b. 2 populatsiooni võitlus ressursi pärast
3. Mutualism (+,+) – kahepoolselt kasulik. Koostöö. Sümbioos.
4. Tarbimine (+,-) – 1 saab kasu teise arvelt
5. Komensialism (+,0) – Nt lagundajad.
6. Amensialism (-,0) – Tehakse tahtmatult halba.
46
1. KONKURENTS (-,-) – kahepoolselt kahjulik
Konkurentsiteooriale pani aluse Gause.
Üksus: populatsioon
Tekib: kui kaks populatsiooni ihaldavad samu ressurse/tingimusi, mida on keskkonnal pakkuda
piiratud hulgal – ressursikonkurents
C1 ja C2 – konkureerivad populatsioonid
E – liikide ühine vaenlane
R – limiteeritud ressurss, mille pärast konkureeritakse
a) VAHETU KONKURENTS
Liikide isendid puutuvad otseselt kokku, otsene võitlus
b) KONKURENTS LÄBI RESURSI TARBIMISE
Kui jaanalind ja antiloop lähevad koos lombist vett jooma, siis jaanalinnu kael on pikem ja
kui lombis jääb vet vähemaks, saab jaanalind juua, antiloop mitte. Võit äbi resursi
ammutamise.
c) NÄIV KONKURENTS
Isendeid ründab sama vaenlane (nt parasiit), kuid ühte mõjutab rohkem kui teist.
nt: Euroopa hirv ja põder – Tarbivad samu ressursse ning hirv lõp põdra alalt välja: Põhjus
ajuparasiidis mis nakatas mõlemat, kuid vaid põder suri.
d) KONKURENTS LÄBI 3.LIIGI
Mitteotsene konkurents, läbi ühise vaenlase, läbi troofilise taseme.
KONKURENTSE VÄLJATÕRJUMISE PRINTSIIP
Sarnaste toitumisharjumustega ligiid on liiga sarnased, et koos elada.
Seadus: 2 liiki, kelle nõudlused limiteeriva ressursi suhtes kattuvad, ei saa pikka aega koos
eksisteerida, vaid 1 neist tõrjutakse süsteemist konkurentselt välja.
Gause konkurentsieksperiment pärmidet toituvate amööbidega
47
Aurelia ja Caundatumi toitumisharjumused on liiga sarnased et koos elada – Caudatum lüüakse
välja.
Caudatum ja Bursaria on piisavalt erinevad, et koos eksisteerida. Nende nišid ei kattu piisavalt,
et üks lööks teise välja, kuid mõlema kasv pidurdub.
E.Hutchinson sõnastas nišiteooriast lähtuvalt Konkurentse väljatõrjumise printsiibi:
- 2 liiki, kelle ökoloogilised nišid kattuvad, ei saa pikka aega koos eksisteerida vaid varem või
hiljem tõrjutakse 1 neist konkurentsist välja.
Limiteeiv sarnasus: Piir, kui sarnased peavad liigid olema, et mitte koos eksisteerida.
Liikide erinev paigutamine ressursigradiendil ennustab erinevat konkurentsiintensiivsust (tänu
erinevalt niššide kattumine määrale).
X-telg – ressursitase/omadus
Y-telg – ressursi kasutamise efektiivsus
w – nišši Laius
Vasakul: nõrk konkurentsi tase Paremal: tugev konkurentsi tase
Paremal: Laiendades nišši amplituudi on 2 võimalust:
Keegi lahkub (keskmine)
Toimub mikroevolutsioon spetsialiseerumise suunas optimum lähedasele ressursile
(mida on kergem kätte saada)
Niššide diferentseerumine – niššide eristumine, tulemuseks stabiilne süsteem
Hutchinson: Planktoni paradoks
Hutchinson vaidlustas ise oma teooria.
On palju nähtusi, mis selle seaduse alla ei mahu. Nt kuidas saab nii palju planktoniliike koos
elada?
Malawi järves söövad sääse vastseid kalad ja paljud muud liigid. Ei saa olla liikide
diferentseerumist – paradox
Paradigma – teadusloos mindite vaadete/mõistete süsteem, mida pole vaja tõestada, võetakse
tõepähe
48
2) MUTUALISM (+,+) – kahepoolselt kasulik
Esineb sagedamini kui konkurents.
Kõikidel hulkraksetel on oma mutualistid.
Mutualism on:
Oblikatoorne – hädavajalik
Fakultatiivne – protokooperatsioon
Mutualismi ligiid:
1. MUTUALISM KUI HÄDAVAJALIK KOHASTUMUS
- Seotud evolutsiooniga
nt: meemäger. Võib sandistada kariloomi. Ründab isaste põllumajandusloomade munandeid ja
sööb ära ning jätab looma surema.
On sõber meenäitajaga (lind). Mõlemaid huvitavad mesilased. Meemäger tahab vastseid ja mett
kuid ei leia taru üles, meenäitaja aga tahab vaha kuid ei saa üksi mesilaste vastu.
Nii leiabki lind pesa, mäger kisub pesa alla ja mõlemad saavad oma osa.
nt: Mosambiigis koostöö põlisrahva ja delfiinide vahel. Põlisrahvale on põhiline kalapüük,
milleks peaks minema ookeani. Kari delfiine ajab aga kalad madalasse vette, kus inimesed
võrguga ootavad. Pool saavad inimesed, pool delfiinid.
nt: Akaatsiad ja sipelgad. Igal akaatsial on oma sipelgas, kes elab akaatsias, akaatsia kasvatab
sipelgatele spetsiaalse organi, pesa. Lisaks kasvatab akaatsia suhkrurikkad väljakasvud, mis on
sipelgatele toiduks. Akaatsiatele on sipelgad aga kaitse putukate ja rohusööjate vastu. Lisaks
lõikavad sipelgad naaberpuude oksad, kui need hakkavad koduakaatsia PARi varjama.
2. PÕLLUMAJANDUS
Tegelevad: inimesed, ühiselulised putukad (termiidid, sipelgad, mesilased)
Inimene ja põllumajandus
Inimene ja kana on mutualistlikud ligiid. Pigem fakultatiivne, ehk mitte hädavajalik. Kodukanu
on 23 miljardit, inimesteta neid nii palju ei oleks. Kasulik liigile ja inimesele.
Sipelgate põllumajandus – 2 liiki
1. lehetäide kari: Lehetäid imevad taimemahlasid selle raku seest. Sipelgad käivad aegajalt
lehetäisid lüpsmas ja annavad sipelgatele tilga suhkruvett. Vastutasuks sipelgad
kaitsevad lehetäisid teiste putukate eest
2. Seenekasvatus – massiliselt tegeletakse sellega, eriti niiskes troopikas. Sipelgad viivad
lehetükke pessa. Nad ei söö lehti, vaid viivad lehed oma seenekasvatusele. Lehelõikajad
sipelgad söövad vaid seeni. Seened muudavad seedimatu biomassi nende jaoks
assimileeritavaks.
3. SEEMNELEVI JA TOLMENDAMISEGA SEOTUD MUTUALISM
On palju loomi ja taimi, kelle eksisteerimine sõltub vaid üksteisest
nt: Loomadel on maasikatega mutualistlik suhe. Masi on neile atraktiivne ja vastutasuks
levitavad liimad maasikate seemneid.
nt: Taimedel on ilusad/lõhnavad õied vaid selleks, et meelitada tolmendajaid. Õie nectar on
mõeldud vaid tolmendajate meelitamiseks, nektarit tarbima tulles jääb putuka külge õietolm.
Ainult koolibri on nekteritoitelised. Iga õie jaoks on oma koolibri. Kui 1 sureb, sureb ka teine.
49
Õietolmu lihtsalt õhku viskamine oleks mõttetu, sest järgmine taim võib olla alles mitme
kilomeetri taga.
4. SÜMBIOOS
- 1 globaalse elu alustalasid.
- pikaajaline intiimne mutualistlik suhe 2 liigi vahel, kellest 1 on alati oluliselt suurema
kehaga kui teine
Selletõttu saab sümbiootiliste suhete põhjal 2 liiki jagada:
1. peremees (suurem)
2. sümbiant
1. KEHA PINNAL
2. KEHA ÕÕNES
3. KEHA ÕÕNES
+
RAKKUDES
4. RAKKUDE SEES
kast – peremeesrakud
ring – sümbiondi rakud
1. Naha pinnal: Inimese nahapinnal on sama palju sümbionte kui rakke – see on kaitse
parasiitsete mikroobide eest. Ilma sümbiontideta läheksime kärna
2. soolestikus: Lõviosa bakteritest meie soolestikus on kasulikud, mis aitavad seedida.
Inimesel puudub tsellulaas, mis mikroobides on olemas ja need aitavad meil taimetoitu
seedida. Inimesel 10 korda rohkem kui keharakke
3. Leidub ainuõõssetel loomadel. Korallidel on see eluviisiks. Korallid elavad sümbioosis
vaguviburvetikaga, kes tegelevad rakkudes ja ainuõõssetes fotosünteesiga – praktiiselt
korallide ainus energiaallikas. Kui korallid paljunevad (suguliselt ja klonaalselt) niin
võtavad nad vetikad kaasa
4. Endosümbiondid! Oli anaeroobne organism, eukarüoot, kes ei talunud hapnikku.
Aeroobsed bakterid tekkisid, sest kasutasid hapnikku elektronide akseptorina. Sellina
bakter neelati endosümbioosi teel amööbi rakku ja jäi seedimata – sündis maailma
esimene mitokonder.
Samasugune endosümbioosi akt pani aluse ka kloroplastidele, kui amööbi neelati
sinivetikad.
50
SÜMBIOOSI ILMING – SEENJUUR EHK MÜKORIISA
Sümbioos taimejuure ja seene vahel. Vaid tänu sellele sai elu väljuda ookeanist ja asuda
maismaale. Maismaa oli enne seda paljas, kalju, mulda polnud.
Senini olid taimed olnud vaid rohevetikad. Rohevetikad olid esimesed taimed, kes maale said.
1. Seened transportisid fosforit ja lämmastikku kaugelt üsna oligotroofses, ehk
toitainetevaeses vees elavale vetikale, kes sai tänu sellele paremini fotosünteesida.
2. Rohevetikad kolisid vaikselt vest välja kaljule, seenjuured olid vees. Sealt said taimed
lisavett ja lisatoitaineid. Sai alguse maapealne fotosüntees.
3. Tasapisi hakkas tekkima muld ja tekkis seenjuur mulla all.
Seenjuur iseloomustab 92% taimeliikidest
Mükoriisa liigid:
1. ENDOMÜKORIISA
Esimesed mükoriisad.
Seeneniitidel ehk hüüfidel on spetsiaalsed organid – arbuskulid,
millega lähevad taime juure sisse.
2. EKSOMÜKORIISA
Puu ehk jämedate juurtega taimede sümbioos seentega.
Seened ainult juure pinnal, mis moodustab juure ümber tupe.
3. ERIKOIDNE MÜKORIISA
Üsna spetsiifiline. Omane vaid sugukonna ericaceae (kanarbiku) esindatatele
nt: pohl, mustikas, sinikas
4. ORHIDOIDNE MÜKORIISA
Sügavalt spetsiifiline. Leidub ainult, ja absoluutselt kõigis, orhideedes.
Orhideed isegi ei idane ilma seeneta, sest tal puuduvad vajalikud toitained. Seene abiga saab
taim toitu, orhidee end ise üksi ära ei elata. Orhideed söövad seda seent, seened on justkui orjad.
(Orhidee ei tee nektarit aga tolmendajad tolmendavad ikka)
Sümbioos, Seotud õhulämmastiku-fikseerivate mikroobidega.
Võime viia lämmastik aminorühma (NH3) koosseisu. See on energeetiliselt väga kulukas.
Mügarbakterid, asustavad liblikõieliste (hernes, ristik, uba) taimede juuri ja moodustavad
juuremügaraid. Nad tegelevad lämmastiku sidumisega, annavad selle taimedele ja saavad vastu
suhkrut.
Liblikõielised seetõttu on suutelised asustama kohti, kus on lämmastiku defitsiit. Seetõttu enne
kartuli kasvatamist kasvatatakse 1 aasta ristikut.
51
Sojaoa juurenoodulid
nt: Lepp – Leppa juurtel asuvad aktinoriissete noodulite kolooniad. Leppa on palju, sest nad on
suutelised kasvama väljakurnatud põllumaale.
Lepa akroniirsed noodulid
nt: Sinivetikas – Sinivetikas Anabaerra azollae on võimega siduma lämmastikku ning ta elab
sümbioosis Azolla filiculoides’iga (sõnajalg) . Kui riisipõllud on vee all, katab vee sõnajalg,
kellega sümbioosis lehtedes sinivetikas. Kui vesi taandub, siis on lämmastikurikas väetis riisile.
52
3. TARBIMINE (+,-)
Üks populatsioon tarbib kas otseselt teist populatsiooni või seda, mida teine populatsioon pakub
(mitte vabatahtlikult)
Tähtsamad nimetused:
1. Kisklus
2. Parasitism (nugimine)
3. Herbivooria
4. Mükotroofia (fungivooria) – seente söömine
5. Jne…
1. Kisklus – Kirjka tapab saagi 1 õnnestunud rünnaku käigus. Tarbib surnuna.
2. Parasiit – On saagiga pikka aega seotud. Eesmärgiks ei ole kunagi saagi tapmine (võib
olla õnnetus). Üldjuhul on üks või mõni saakloom. Konsurmeerib organeid aga jätab
saaklooma ellu
Parasiidid + parasitoidid – 10% maakera liikidest. Parasiidid munevad munad
saakobjekti (enamasti vastse) sisse (enamus putukad).
Nt: herilane muneb liblika nuku sisse, liblika asemel väljub nukust herilasi.
3. Toit on paigal, tarbja liigub
Strateegia, mis on omane herbivooridele ja pmst kõikigele – NÄKITSEJA. Näkitseja tarbib
tükikesi saagist.
Nt: Gomodo varaan (maa suurim sisalik) – kiskja rebib kitselt tüki küljest, seejärel jälitab, kuni
sureb ja tarbib ülejäänu.
Nt: Põder näkitseb pajupõõsast.
LETKA-VOLTERRA TÜÜPI KISKJA-SAAKLOOM VÕRRANDSÜSTEEM
Matemaatikud modelleerisid diferentsiaalsüsteemi abil bioloogilisi interaktsioone.
Üksiku popultasiooni piiratud kasvuvõrrand:
dN/dt = rN ( 1 – N/K)
rN: paljunemisest tekkiv kasv (pidurdav takistus)
1 – N/K: liigisisene konkurents
Saaklooma puhul on unustatud 1-N/K, sest populatsiooni suurust kontollib eelkõige kirkja.
Top-down control: saaklooma populatsioon on kontrollitud saakloomade poolt. (troofilised
tasemed)
N - saaklooma populatsiooni tihedus
P - kiskja populatsiooni tihedus
r - võime ajaühikus järglasi saada
a’- toiduotsingu efektiivsus . Liigiomane koefitsent. On mõlemapoolne!
fa’NP – mis saakloomale halb, kiskjale hea
f - kiskja liigiomane koefitsent. Efektiivsus millega tarbitud toit konveeritakse järglasteks.
(tiger sõõb hunnikutes liha ja aastas kaks poega vs boa ei kiirga kogu aeg FR tagasi ja saab
aastas sadu järglasi)
qP – kiskja jaoks peamine takistus, liigisisene konkurents, mis sõltub tihedusest.
dN/dt = rN – a’NP
dP/dt = fa’NP – QP
53
Kui mõlema populatsiooni muutumiskiirus on 0, siis on võrrandisüsteem tasakaalus – on
stabiilne.
dN/dt = 0 => rN = a’NP => r = a’P => P=r/a’
dP/dt = 0 => fa’NP = qP => fa’N = q => N=q/fa’
Stabiilne olek – punkt, mis rahuldab mõlemat võrrandit.
Üldjuhul süsteemid sinna punkti ei jõua.
P
r/a’
q/fa’ N
Kui jänesed ja kiskjad saavad 1. Korda kokku, ei hakka kõik liikuma tasakaaluasendi poole,
vaid tasakaal käib mööda ebaühtlast rada.
SAAKLOOM KISKJA
Mida pikem vektor, seda tugevam tõmme.
Nt: Süüa vähe, langeb ruttu.
SUMMAARNE FAASIPORTREE – kujutab süsteemi muutumist ajas, puudub ajatelg.
Oranž – resultant. Summavektor. Näitab süsteemi liikumissuunda
Süsteem hakkab käima ringiratast ümber fookuspunkti
Ringliikumine väljavenitatuna annab sinusoidi, kus veerandperioodilise vahega
liiguvad, kiskja ees, saakloom järel.
Stabiilse seisundi
tingimused. Selline
tihedus, mille puhul on
mõlemad populatsioonid
korraga nullkasvus
54
Peremehe ja parasitoidi dünaamika
PARASITISM
Üle 50% maakera liikidest on parasiidid.
Seni pole leitud ühtegi hulkrakset organismi, kellel poleks liigispetsiifilisi parasite, kellel
omakorda on liigispetsiifilised parasiidid.
• Mikroparasiidid – ei suuda loendada, saab määrata nakatunud peremeeste järgi.
• Makroparasiidid – saab loendada. Nt puuk
• Hemparasiidid – taimed: poolparasiidid. roheliste lihtedega taimed. Fotosünteesivad +
varastavad naabertaimede juurtest aineid. Nt: robirohi, silmarohi.
• Holoparasiidid – taimed: täisparasiidid. rohelisi lehti pole.
Enamik haigusi on mikroparasiitide põhjustatud (viirused jne)
Millest sõltub parasiitide levimiskiirus?
R0 – puhas kasvukiirus. Mitu järglast keskmiselt populatsiooni liige elu jooksul toodab. Kui
mitu nakatunut 1 populatsiooni liige elu jooksul tuudab
R0 = 1 – epideemiat pole, ei levi, ei taandu
R0 < 0 rünnak taandub
R0 > 0 epideemia levib
Mis määrab R0?
R0 = LSB
L – aeg, mille jooksul peremees jääb nakatunuks. Mida pikem L, seda rohkem jõuab levida
S – nakkusele vastuvõtlike isendite tihedus (kel puudub immuunsus, kaasasündinud v tekitatud)
B – mikroparasiidi ülekande kiirus, virulentsus. Kui kiiresti parasiit levib.
Leetrid levivad tohutult kiiresti. AIDsi on tunduvlt raskem saada.
Immuniseerimise eesmärk: R0=1=LSB
=> S(im)=1/LB
Populatsiooni
hulk,
mis
tuleb
immuniseerida,
et
epideemia
ei
leviks/epideemia tõkestamiseks.
P = 1 – S(im) / S(s0)
S(s0) – algne populatsiooni tihedus.
Mis % on vaja immuniseerida, et taanduks. ->
55
SAAKORGANISMIDE KAITSEKOHASTUMUSED
1. GENEETILINE KAITSE – odavaim, evolutsioonis kergeim tekkima
Mürgised on maailmas paljud ligiid (ka mikroobid, seened, protistid ja loomad).Selliseid
liike saavad tarvitada vaid need, kellel on kohastumus – vastumürk. Kunagi pole 1 liik
mürgine kõigile
2. MEHHAANILINE KAITSE – Iseloomulik paljudele hulkraksetele aga ka protistidele
(ränivetikate ränist kest, kilpkonn, karbid, siil, roosi okkad)
3. KÄITUMUSLIK KOHASTUMUS – Vajab intellekti, et leida endale turvaline peapaik
(urud, kaljupraod) millele on lisaks sotsiaalne koostöö vaenlase märkamiseks
Nt: surikaadidel on väljas valvurid
4. VÄRVUSEGA SEOTUD KOHASTUMUS – nii värv kui muster. Kohastumuste skaala
on lai.
Varjevärvus – mitte eristumine substraadist. Nt putukad kes näevad välja kui puulehed.
Kameelion – suudab värvi muuta
5. EHMATAV VÄRVUS – Paljudel liblikatel. Nt päevapaabusilm. Tema tiibade alumina
pool on substraadi värvi, kui keegi tuleb teda ründama siis ta arvab tiivad, ning mustad
ringid tunduvad kui silmad.
Samuti ehmatatakse kujuga.
6. SEGAV VÄRVUS – nt sebra. Kui lõbi ründab sebrasi, siis joostes tekib triipude sigrimigri
– ajab lõvi segadusse.
7. MIMIKRI – mõne teise organismi jäljendamine.
Mimikri 2 tüüpi:
1. BATESI MIMIKRI – Kedagi jäljendades valetad, pettuse eesmärgil.
Nt: Õiekärbes on täiesti kahjutu, herilase triipudega. Seetõttu teda kardetakse.
Nt: orhideedel on õied ainult selleks, et tolmendajaid saada.
See ei saa kunagi saada valdavaks – kaitse kaoks
2. MILLERI MIMIKRI – Mürgised/mittesöödavad organismid jäljendavad üksteist.
Signaali tekitamine ja võimendamine.
Nt: väga mürgised väikesed konnad on kõik tohutult värvikirevad.
Batesi mimikri nõrgestab Mulleri mimikrit. Mulleri mimikri toidab Batesi mimikrit.
56
4. KOMENSIALISM (+,0)
Tarbitav kahju ei saa, tarbja saab kasu.
Mullas on laguahel – orgaanilise aine mineralisatsioon, aine dissimilatsioon. Ilma laguahelata
puuduks orgaaniline aine.
Laguahela produktid: CO2, H2O, muud orgaanilised ained
SAPROTROOF – Surnud, orgaanilisest ainest toituja, lagundaja. Ei tee kahju sellele, keda
lagundavad.
Laguahel on doonorkontrollitud süsteem – nad ei tapa kedagi, ei mõjuta otseselt toiduobjekti
tihedust. Ka raisakotkas ei tapa, kuigi eeldused on olemas.
Tohutult vähe erandeid: nt nekrotroofsed parasiisis – seened. Need on männi okkaid nakatavad
seened, käituvad parasiidina, kuni männiokas elab. Kui männiokas sureb, söövad surnud okast
edasi.
HOOGHÄNNALISED – meie kliimas olulised.
Mikroobsed lagundajad saavad kasu suurematelt lagundajatelt enne neid – kakandite kohalolu
kiirendab oluliselt lehevarise mikroobset lagundamist. (neid ei tohi olla ka liiga palju)
Erinevate orgaaniliste ainete lagunemiskõverad tammemetsa varises. Tselluloos laguneb hästi,
seentel tsellulaasi palju. Palju raskem on ligniini lagundamine.
57
Protsesside intensiivsus sõltub, millise bioomiga on tegu. Mikro- meso- ja makrofauna
aktiivsus laguahelas eri bioomides. Allpool toodud ribade Laius märgib varies lagunemise
kiirust ja rundud orgaanilise aine akumulatsiooni kiirust.
Mikrofauna: bakterid
Mesofauna: hooghännalised, väikesed putukad
Makrofauna: vihmauss, silmaga nähtav.
Meie asume mesofauna tipus.
1m3 parasvöötme (Eesti kuuse-sega) metsa.
1000 loomaliiki (lagundajate hulgasi, seened/bakterid välja)
10 000 000 protisti ja neumatoodi
100 000 hooghännalist ja lesta
50 000 muud selgroogset.
Austraalia
1788 oli Austraalias 7 lehma.
1960 oli 30 000 000 lehma.
1 lehm = 10 kooki päevad. Kokku 300 000 000 kooki päevas.
See katab 3 000 000 hektarit. Kohalikud lagundajad olid harjunud känguru sitaga – ei
lagundanud. Inimesed kaotasid karjamaid.
1963: inprodutseeriti sinna Aafrikast 21 liiki sõnnikumardikaid, 2 neist jäid ellu – inimesed said
karjamaa tagasi.
Interaktsioonide lõpp.
58
SÜNÖKOLOOGIA – looduse koosluste uurimine, mis on tervikuna pea võimatu.
Koosluste struktuur:
• Vertikaalne (taimed koosnevad rinnetest ehk taimkatte kihtidest, preerias pole aga
metsas pea alati on)
• Horisontaalne (koosluste muutumine keskkonna gradiendil. Igal gradiendil oma
liigikomplekt.
• Bioloogiline mitmekesisus – elurikkus, diversiteet. Antarktikas kasvab 2 liiki taimi,
Amazonases samal pindalal kasvab tohutult rohkem.
• Taksonoomiline strukruur – Suktsessioon – koosluste muutumine ajas (suunatud, vahel
korduv). Koosluste ajaline arend nii iseeneslikult kui väliste jõudude põhjustatult.
Sünökoloogia algas eelmisel sajandil.
Koosluste energertsed vs kollektiivsed omadused – suur vaidlus
Kas kooslused on diskreetsed (mitte pidevad) koevolutsioneerunud üksused, süsteemid, kus
liigid on üksteisega pikka aega kohastunud (kaitse/koostöö) või on tegemist konglomeraadiga,
mille omadused määravad liigid, kes sinna kokku on sattunud?
CLEMENTS: Sünergeetilist efekti väljendav omadus. Komponendid koos tekitavad uusi
omadusi, mida komponentidel endal pole (pannkook). Liigid diskreetsed. Ei saa ainult isiklikult
komponentide kaudu kirjeldada kooslust, vaid nad on koevolutsioneerunud süsteemid. Liigid
on üksteisega kohanenud ja sõltuvad üksteisest.
Clementsi ideed: Viisid koosluste klassifitseerimiseni
GLEASON: Individualistlik. Kooslus pole tervik, vaid pigem seda moodustavate liikide
omadused määravad lihtsalt selle, kes seal keskkonnas hakkama saab. Liigid käituvad
keskkonna järgi, ei moodusta süsteemi.
Gleasoni ideed: Viisid ardenatsioonini. Liike saab järjestada/võrrelda sarnasuse alusel.
1. Koevulutsioneerunud süsteem – organistlik paradigma CLEMENTS – Euroopa
Clements vaatles taimekooslusi kui diskretseid ja koevolutsioneerunud süsteeme, millel
on algus ja lõpp.
2. Liikide konsfomeraat - Individualistlik paradigma GLEASON – Ameerika.
Mõlemal on natuke õigus, mõlemad natuke eksisid.
Gleasoni maailmapilt oli ökoloogiale nurgakiviks.
Preerias diskretseid kooslusi ei leiagi.
59
Eestis on tugev metsateaduste koolkond.
1 kooslustüüp: metsa kasvuala tüüp.
Erich Lõhmas: Koostas klassifitseeritud süsteemi
Vasakult-paremale: mulla viljakus
Ülevalt-alla: üldistatud niiskus
KOOSLUSTE SUKTSESSIOON – Koosluste areng (sisemistel/välimistel põhjustel) /
välimine muutumine
Jaguneb:
1. PRIMAARNE suktsessioon – Toimub seni taimestumata pinna. Nt: mere alt
vabanenud aladel. Uued tekkivad vulkaadilised saared.
Nt: Põhja-Ameerikas liustike alt vabanev kivine maa.
2. SEKUNDAARNE suktsessioon – Järgneb peale mingit suktsessiooni häiringut. Väliste
jõudude poolt tingitud bioloogilise massi suremus/massiline ärakanne.
Häiringufaktorid: nt tulekahju, torm, üleujutus, massiline parasiitide/herbivooride
rünnak, metsaraie jne.
Nt: raiesmikust areng küpse metsa suunas.
Primaarne suktsessioon: Põhja-Ameerikas liustike alt vabanev kivine maa.
Logaritmiline ajatelg.
Lepp (alder) elab koos mikroobiga perekonnast
frankia, kes suudab siduda lämmastikku.
60
Clements püstitas teooria: Iga keskkonna jaoks on olemas oma suktsessiooni lõppstaadium
ehk kliimakooslus, mis on jääv.
Kliimakooslus-kliimaomane suktsessioon. Lõppstaadium, mus ei muutu häiringuteta miski.
See on monokliimaksi teooria. Eestis on koosluseks peetud kuuse-segametsa, välja arvatud
liivaluiteid, kus kasvavad männid. Lisaks ka kõrgsoo ehk raba.
Kliimaks – kliimale omase suktsessiooni lõppstaadium. Kui kliima ei muutu.
SOODE SUKTSESSIOON.
Soo areneb mitmes faasis, tihti madalate veekogude
kinnikasvamisel.
1. MADALSOO
Taimede juures ulatuvad mineraalsesse mulda. Lagundajad
ei saa seisvas keskkonnas hakkama, sest seisvas vees on
hapniku defitsiit. Anaeroobsed ei saa kunagi lõpuni
lagundatud – hakkab tekkima turvas.
Turvas koosneb lagundamata taimejäänustest, ehk peamiselt
on seal süsinikku, lämmastikku ja fosforit pole. Väga
toitainetevaene.
2. SIIRDESOO
Produktsioon toimub edasi, kuni lõpuks taimedel juures ei
ulatu enam mineraalmulda, vaid on turba sees.
Tekkivad oligotroofsed mättad – hakkab kasvama
teistsugune taimestik.
Tekib siirdesoo. Siirdesoos asuvad oligotroofsed mättad.
Hakkab kasvama turbasammal.
3. KÕRGSOO
Taimed kasvavad ainult turbal. Kõik on turbasamblaga
kaetud.
10% taimedest jääb soos alati lagundamata – nii tekib Eestis ca 1mm aastas turvast juurde.
Raba on ombrgiotroofsed – vihmavee toitelised. Kõik toitained saavad rabataimed vihmaveest
– seetõttu ongi kidura kasvuga, aga elavad.
Raba tekib kui keskkonnas on:
1. Liigniiskus
2. Seisev vesi – vesi ei rikastu hapnikuga
Ökosüsteem, kus on turvas olemas – SOOSTUV süsteem. Seal, kus turbasammal on õhem kui
30 cm. Nt: soostuvad metsad
Turbasammal on nii lämmastikuvaene, et ei ahvatle herbivooridele.
61
BIOLOOGILINE MITMEKESISUS EHK DIVERSITEET
Koosluses kooselavate bioloogiliste liikide arv. Mida rohkem on liike, seda mitmekesisem on
ökosüsteem.
Liikide lugemisele lisaks võib lugeda ka teisi teksoome. Eriti nt fossiilide määramisel, ei saa
tihti liiki määrata, vaid saab määrata nt sugukonna.
Kooseksisteerivate liikide arv sõltub: eelkõige evolutsoonist.
Diversity hotspots – keskmisega võrreldes on palju rohkem liike. Täpselt seletust pole, mis
kust nt Andides on diversity hotspot.
Bioloogilised liigid maakeral: 1,75 – 1,9 miljonit (20% neist võib olla sünonüüme) on
dokumenteeritud.
Liigirikkus taksonite vahel jaotub ebaühtlaselt.
Nt:
1000 000 lülijalgset, 400 000 mardikat
Nt:
310 000 taime: 282 000 soontaime, 100 000 seent.
Liigirikkuse rekordid (pindala kohta):
Domineerivad 2 sorti kooslust:
Euroopas – parasvöötme niidud + rohumaad
Lõuna-Ameerika – Troopiline vihmamets
Laelatu puisniit Eestis – praktiliselt iga taim erinev 0,1 m2 kohta.
Liigirikkuse seotus laiuskraadiga.
Ekvaatori lähedal on liigirikkus suurim.
Erandeid on vähe:
1. Samblikud ehk jägalad – lihhemiseerunud seened. Vetikad seal fotosünteesivad.
2. Avamerelinnud – kes pesitsemise välisel ajal avamerel palju külmades meredes
(pingviin)
62
Sarnane seaduspära on ka:
1. absoluutse kõrgusega: kõrgemal on vähem organisme, liike
2. Sügavuse gradient: umbes 1km sügavusel liigirikkus suurim. Üleval vähe toitaineid,
allpool fotosüntees raskendatud.
Liigirikkus a) laiuskraadi ning b) absoluutse kõrguse gradiendil.
a)
b)
Liigirikkus sügavuse gradiendil
Endeemsed – omased vaid sellele keskkonnale.
Vaadeldes liikide arvu, on maismaal palju rohkem liike. Maa on struktureeritum üksis kus
kohastumusvõimalusi on rohkem.
63
LOKAALSED DIVERSITEEDIGRADIENDID
1. POOLSAARE JA LAHE EFFEKT – TÕENÄOSUSLIK SEADUSPÄRA
Lahe tipu suunas diversiteet väheneb. Poolsaare tipus/lahe otsas on mitmekesisus väiksem.
Mereliigid tekivad meres, ookeanid ja lahe tippu tuleb neil migreeruda. Liigid migreerivad igas
suunas ja lahesoppi sattumine on ebatõenäoline. Samamoodi on mandril.
Nt: Lahe effekt Läänemeres. Ligipääs ookeanist on vaid läbi Taani väinade. NB! Lisaks mängib
rolli ka siin soolsus. Kaugus ookeanist korreleerub soolsuse vähenemisega.
2. NN KÜÜRSELG KÕVER (hump-back curve)
- Nime andis P.Grinne
Küürselg kõver sidus produktsiooni läbi biomassi liigirikkusega.
Seos on unimodaalne, vastandub monotoonsele seosele (mis ainult kasvab/kahaneb mööda x-
telge).
Enim liike kasvab mõõdukalt vaestel muldadel –
produktiivsuse tõustes läks liigirikkus alla.
On olemas vähe liike, mis on kohastunud
keskkonnale, mis on ebasoodne. Kui muld on
mõõdukalt vaene: kõik kes tulevad, jäävad ellu. Edasi
liigirikkus langeb, sest intensiivistub liikidevaheline
konkurents. Taimed kasvavad siire,als. Liogo
viljakat mulda armastavaid taimi on palju, jäävad ellu
tugevamad.
64
LIIGIRIKKUS S VS PINDALA
Liigirikkuse ja pindala vahel on lineaarne seos.
Liikide arv (S) on pindalaga seotud läbi 2 konstandi:
S = cAz
Log S = log c + z log A
z – näitab seose intensiivsust, ehk tõusu (nii vees, kui maismaal)
saartel: suur z
mandril: väike z
Seda põhjustab levimine. Saarele migreerumine ja seal ellu väämine on keeruline. See teeb
saare suuruse kriitiliseks. Kunkurents väikesel saarel on suur, liike on väha ja võib ette tulla
palju õnnetusi. (saarel on 3 kitse, kogemata tapad kõik).
Globaalne liigirikkus evolutsioonilises ajas
– positiivse tagasiside põhimõttel. Mida rohkem liike, seda rohkem liike.
Evolutsiooniline aeg alates kambriumist-tänapäevani.
LOOMAD
y-telg – fossiilsete selgsoogsete seltsiline mitmekesisus.
Selgroogsete arv: peaaegu eksponentsiaalne.
Algab: kahepaiksetega, hiilgeaeg permis, edasi
suhteliselt stabiilne.
Roomajad: suur tagasilöök seotud meteoriidiga
Imetajad: liigirikkuse tõus plahvatuslikult
Linnud: samuti plahvatuslik kasv
Luukalad: samuti plahvatuslik kasv sarnaselt
imetajatele ja lindudele
TAIMED
Kui loomariigis on justkui kumulatiivne, siis taimeriigis
on toimunud pigem asendumine.
a) Ossid
b) sõnajalgtaimed (max kambriumis - tekitas kivisöe)
c) paljasseemnetaimed
d) katteseemnetaimed – kriidis, kui dinosaurused välja
surid.
65
Põhjus: kõik taimed ihkavad samu ressursse. Nt metsvindi ja arusisaliku toidulaud on täiesti
erinev, toimub pidev spetsialiseerumine uutele ressurssidele.
Mass-ekstinktsioonid – tagasilöögid.
Üldist väljaduremise kiirust iseloomustab must paks joon – languses, sest liigid on muutunud
võimekamaks.
a) 1. langus: perm
2. langus: triias (meteoriit) - sai saatuslikuks suurtele organismidele ja
veeorganismidele.
66
EVOLUTSIOONILINE ÖKOLOOGIA
- Seletatakse evolutsiooni läbi ökoloogiiste protsesside.
- 1 suur põhiküsimus: miks?
- Vaatluse all on elukäigutumus – elulised/olulised omadused, mis on seotud organismi
kasvu/paljunemisega.
Nt: keha suurus, eluiga, paljunemispingutuste arv, järglaste arv, ....
- Eesmärk selgitada, kuidas evolutsioon võõi elukäigutunnusteni.
Evolutsiooniteooriale panid aluse Darwin ja Wallace.
Optimaalse kurna suuruse kujunemine (clutch size).
Järglaste arv 1 paljunemispingutuse käigus.
a)
Evolutsioon liigub alati suurema fitnessi suunas. Suurem fitness = rohkem elujõulisi järglasti
(lapselapsi). See on genotüübi suhteline ppanud tulevastesse põlvkondadesse.
Mida suurem kurn, seda suurem fitness
b)
Olemas optimum, kus summaarne kohasus on suurim.
Linnud on iteropaarsed – palju paljunemispingutusi (vs iteropaarsed – 1 paljunemine elus).
Väga suur kurna suurus kurnab emast ja on võimalus, et ta ei saagi rohkem järglasi.
Enese säilitamine on oluline!
c)
Paljunemine hind – tuleb kontrolli all hoida.
Võimalikult suur summaarne kohasus järglastele ja ema säilitus.
Koht, kus optimumaalse kunra suuruse vahe on suurim.
67
LÕIVSUHE
- Negatiivne seos erinevate võimekuste vahel piiratud ressursi tingimustes
- Seos, mis tuleneb lõpliku hulga ressurside ümber paigutamisest erinevate ressursside vahel
Kuidas toimub lõivsuhe?
Allomeetrilised seosed – Positiivne seos, nt kaal ja käte pikkus.
Lõivsuhe pole paratamatu, väike ploomipuu – vähe lapsi.
Varieeruvus
lookuses
A
tekitav
varieeruvuse
ressursihõlvamise edukuses ja viib allomeetsilise seose
tekkimisele (kollased täpid all graafikul). -Paremini
ressurssi haaravad isendid kasvavad paremini ja
paljunevad rohkem.
Geneeitline
varieeruvus
lookuses
A
–
viib
allomeetrilisele seosele: mida paremini omastab seda
paremini saab.
Varieeruvus lookuses B tekitab varieeruvuse ressursside
allokatsioonis ühelt poolt kasvu ja elutegevusse, teiselt
poolt paljunemisesse. Selline varieeruvus avaldub
lõivsuhtena (mustad täpid graafikul)
Geneetiline varieeruvus lookuses B – tekitab lõivsuhte:
palju suurem karv = paljuned halvemini jne.
1. Ressursid
2. geenilookus 2 alleeliga
3. (allopatsioon – ümberpaigutamine. Valiku küsimus, harva teadlik)
4. a) ressursid suunatud ellujäämisesse või b) ressursid paljunemisesse suunatud
Tänu sellele valikule tekivki lõivsuhe.
Nt: Käbide arv puu kohta vs aastane kasv – lõivsuhe ebatsuuga
käbitoodandu ja aastarõnga laiuse vahel
Nt: Lõovsuhe seos isase puuviljakärbse suuruse ja
eluea vahel (suuremad elavad kauem) peidab endas
lõivsuhet isase seksuaalse aktiivsuse ja eluea vahel.
Täpid näitavad isaseid, kes pandi kokku elama
kaheksa viljastatud emasega. Numbes 1 on tähistatud
isased, kes said kaaslaseks ühe viljastamata emase ja
seitse viljastumata emast ja nr 8ga tähistatud isased
elasid koos kaheksa viljastamata emasega. Nende
eluiga oli 2 korda lühem. Suguline aktiivsus lühendab
eluiga.
Lõivsuhe ja allomeetriline seos.
Varieeruvus lookuses A tekitab
varieeruvuse ressursihõlvamise
edukuses ja viib allomeetrilise
seose tekkimisele (kollased
täpid alumisel graafikul) –
paremini resurssi haaravad
isendid kasvavad paremini ja
paljunevad rohkem.
Varieerumine lookuses B
tekitab varieeruvuse
ressursside allokatsioonis ühelt
poolt kasvu ja elutegevusse,
teiselt poolt paljunemisse.
Selline varieeruvus avaldub
lõivsuhtena (mustad täpid
alumisel graafikul).
68
SEKS
Kõrvuti loodusliku valikuga eksisteerib looduses suguline valik. Suguline valik ei pruugi aga
olla kohasust suurendav.
Seks kinnistus evolutsioonis kiiresti. Sugulise paljunemise eelised?
- Reaalselt paljunevad vaid pooled emased, klonaalselt kõik.
- Toodetakse lõpmatutes kogustes uuri genotüüpe – rohkem materjali looduslikuks valikuks.
Nt: keskkonnatingimute muutudes on võimalik kiire evolutsioneerumine. Klonaalselt
paljunevad peavad ootama somaalset mutatsiooni
Soo määramise mehhanismid
1. Sugukromosoomid (inimesel)
2. Haudumise temperatuus (roomajatel)
3. Viljastamine (ühiselulistel putukatel: viljastamata – isane, viljastatud – emane)
Sugu võib elu jooksul vahetuda (nt lõhe).
Kaladel vahetub sugu elu poole pealt.
Soovahetuse mõistlikkus ja ajastatud sõltub paljunemisedukuse ja kehasuuruse seosest emastel
ning isastel
a) Soovahetust ei toimu
b) Suurem isane saab paljuneda, väiksem ei saa. Seega on mõistlik olla elu alguse poole
emane.
c) Kui isased ei võitle. Mõttekas on olla alguses isane (saab rohkem viljastada) ja hiljem
emane (suurem ema, rohkem lapsi).
Proandria – noorena isane
Protogüüria - noorena emane
Hermatradism: olemas nii emas kui isas suguorganid. Nt: taimed, kus õied on
mitmesugulised. (kadakad on nt lahksugulised)
Sugu selgub alles paaritumise käigus. Enamus maakera taimedest on lahksugulised.
FISHERI REEGEL: Sugusid sünnid alati võrdselt.
1 erand: ühiselulised putukas, kus sugu saab valiga.
Evolutsioon soodustab alati protsesse, et isaseid ja emaseid oleks võrdselt. Ja kui mingi sugu
hakkab väheusse kalduma, siis kisub evolutsioon sinna poole. Evolutsioon soosib alati
vähemuses sugu.
Nt: Olgu 6 naist ja 3 mees – tuleb 6 last. Naise kohasus: 1 laps naise kohta , meeste kohasus: 3
last mehe kohta. Seega on meestel suurem kohasus.
69
Monogaamia vs polügaamia
Monogaamia – 1 püsiv partner.
Imetajad, tihti linnud. Roomajatel haruldane, ainus monogaamiat harrastav roomaja on
käbiloom (sisalik)
Polügaamia – mitu partnerit elu jooksul
Eriti levinud seriaalne polügaamia: igal paljunemispingutusel on uus partner
Suguline valik: võistlus parema partneri üle, lõpliku valiku teeb emane. Selleks on tunnused,
mille järgi emased saaksid otsustada isaste kvaliteedi üle. Suguline valik toimub ka
sugurakkude tasemel.
Suguline dimorfis: äratuntavalt peale vaadatavalt morfoloogilised erinevused isaste ja emaste
vahel – selleni on viinud suguline valik.
Sugulise valiku tasemed
1) Suguraku tase
2) Võitlus (nt Ameerika piisonid)
3) Händikäp-käitumine, händikäp tunnus.
Teatavad tunnused näitavad ausalt isase head kvaliteeti ja tervist, mis organismile endale on
aga kahjulikud. Enesele kahjulik käitumine/tunnus, eesmärgiga näidata oma head kvaliteeti.
Nt: isase paabulinnu saba. Paabulinnul on nii hea tervis, et ta saab endale lubada suurt saba.
Kuigi seda on raske vedada, see paistab röövloomadele silma, on tal õnnestunud ellu jääda.
Järelikult on tal head geenid
Nt: lõvi lakk. 20h ta ööpäevas magab, sest lakk on palav ja kihiseb parasiitidest. Aga ta saab
hakkama.
Nt: inimestel toimub suguline valik. Kamba liidrid saavad ilusaimad emased. Ka ilus auto, kui
oled ostnud enda raha eest, on eelis. Meeste käitumine on seotud sugulise valikuga
Nt: Lehtlalindude isased ehitavad lehtlaid, kus toimub vaid paaritumine. Meeste ainus ülesanne
elus on lehtlate ehitamine.
nt: Linnu saba eksperimetnaalse lühendamise või pikendamisega
saab mõjutada linnu ek’dukust partneri leidmisel. Emased
eelistasid suurema sabaga linde, keda looduses ei eksisteerigi
4) Võistlus
Isased esinevad ilma agressiivseks muutumata. Nt: metsis, teder. Mida parem tevis, seda
paremini ja rohkem jõuad esineda
Nt: Isalindude etteastete arv päevad vs sellele kulutatud
energia. Ringiga – paaritusid.
70
5) Geneetiline valik.
Valik reeglina lõhna järgi või muude eritiste jätgi paaritumisel. Lenduvad keemilised ained,
mis annavad aimu geneetilisest sobivusest.
Feromoonid. Emased on neile väga tundlikud. Putukate emased võivad õige isase kilomeetrite
tagant ära tunda. Ka inimestele on see oluline. T-shirt test.
71
AINE- JA ENERGIA LIIKUMINE ÖKOSÜSTEEMIS
Me assimileerime nii ainet kui energiat.
PRODUKTIIVSUS: kui palju salvestatakse primaarproduktsiooni käigus ehk kui palju on
ökosüsteemis ainet/energiat.
Puhas primaas NPP = GPP (gross primary production) – R (hingamine)
Ökosüsteemide produktiivsus maismaal ja ookeanis
Ökosüsteemide primaarset produktiivsust mõõdetakse kas energia või aine (nt C)
salvestamisena pindala kohta ajaühikus. Koguproduktsiooni tähistatakse inglise keelse
lühendina GPP, ehk taimede enda elutegevuseks kuluv osa, saame puhta primaarproduktsiooni
NPP. Maismaal seletub produktiivsuse muster üsna hästi aasta keskmise temperatuuri ja
sademete hulgaga. Allpool olevast tabelist on näha, et kõige produktiivsemates
maismaaökosüsteemised on NPP kahe kilogrammi kuivaine kandis ruutmeetril aastas, meie
parasvöötmemetsades on ca kaks korda vähem.
Maailmameres pole sademetel mõistagi mingit rolli, samuti on aasta keskmisel temperatuuril
vähe tähtsust. Produktsioon koondub ookeanid peamiselt mandrilavade lähedusse, kud
toitaineid pidevalt ülemistesse veekihtidesse uutakse. Ookeanide keskosad on aga kõrbed, kus
produktsioon on minimaalne. Selle põhjuseks on toitainete akuutne defitsiit (eelkõige forfot ja
raud). Ookeanide keskosad on sügavad ja põhjakihtidest satub mineraalseid toitaineid
pinnakihtidesse väga harva.
Pööra tähelepanu kahel maakaardil olevate tingmärkide absoluutsele skaalale, maismaa mõttes
on tegelikult ookeanid kõrbed.
72
Energia ja aine ülekandumine – ökoloogiline püramiid
Kümnendik 1 troofilisest tasemel assimileeritud energiast kandub edasi järgmisele tasemele.
Ülejäänu läheb DOMi (söömata jäänud osa) – laguahelasse. Lisaks kaob energiat respiratsiooni
teel maailma.
Troofilised tasemed:
1. Taimed
2. Rohusööjad
3. Rohusööjate sööjad
4. Tippkiskjad, söövad ka kiskjaid.
Energia liigub mööda toiduvõrgustikku: 1 pall = 1 liik. Toiduvõrgustik on läbipõimunud ja
keeruline
Energia liikumine ühelt troofiliselt tasemelt teisele
Allpool näha olev skeem illustreerib energia ülekannet troofiliste tasemete vahel. Skeem
kujutab torustikku, kus energia liikumise alternatiivsed rajad on eri torudes ja viimaste
läbimõõte peegeldab “vooluhulka” vastavas torus.
Pn ja Pn-1 on koguproduktsioon n-dal ja sellele eelneval troofilsel tasemel. Näeme, et alumisel
tasemel salvestatud energiast läheb suur osa otse laguahelasse, ülejäänu on sisendiks (In) n-da
troofilise taseme jaoks. Kogu sisendit (sisse söödud orgaanilist päritolu energiat) ei suudeta
mõistagi assimileerida (seedida). Osa väljutatakse fekaalse kaona Fn, mis läheb laguahelasse.
73
Seeditud osa moodustab n-dal tasemel assimileeritud energia An. Kogu assimileeritud energiat
ei saa kasutada produktsiooniks, sest osa kulub organismide elutegevuseks e. hingamiseks Rn.
Mis sellest üle jääb, moodustab produktsiooni n-dal troofilisel tasemel Pn. Energia liikumise
erinevate etappide kohta saame välja rehkendada mitmesuguseid efektiivsusi (kui tahame
viimaseid protsentides tuleb need 100%-ga korrutada):
Tarbimisefektiivsus CE = In/Pn-1 – näitab kui efektiivselt suudetakse eelmisel troofilisel
tasemel salvestatud energiat tarbida;
Assimilatsiooniefektiivsus AE = An/In – näitab, millise efektiivsusega suudetakse tarbitud toitu
assimileerida;
Produktsiooniefektiivsus PE = Pn/An – näitab, milline osa assimileeritud energiast suudetakse
produktsiooniks pöörata.
Kui me toodud efektiivsuses korrutame, saame energia ülekande efektiivsuse kahe troofilise
taseme vahel TE = CE x AE x PE = Pn/Pn-1.
Allpool oleval skeemil on näidatud energia liikumise efektiivsused erinevate taksonoomiliste
loomarühmade ja erineval troofilisel tasemel paiknevate loomade jaoks. Palun süvenege
skeemidesse ja mõelge välja, mis üks või teine efektiivsus on tähelepanuväärselt madal või
kõrge.
74
GLOBAALSED AINERINGED
Aineringete puhul huvitavad meid ennekõike peamised fondid (i.k.pools) ja vood (i.k.fluxes).
Esimene näitab, kus mingi aine või element püsivalt paikneb, teine näitab, milliste fondide
vahel aine või element ümber paikneb. Fondide suurust mõõdetakse massiühikutes, voogude
suurust aine ümberpaiknemise kiiruse ühikutes (massiühikut ajaühikus).
1. SÜSINIKURINGE
Allpool olevalt skeemilt näeme, et maakeral paikneb ringes olev süsinik peamiselt neljas suures
fondis (suuruse järgi reastatuna) – ookeani fond (38000 ühikut), mulla fond (1500 ühikut,
sisaldab taimejuuri), atmosfääri fond (720 ühikut), maismaataimede maapealse osa fond (560
ühikut). Lisaks sellele on süsinikku mõistagi massiliselt ka igal pool mujal (fossiilsed kütused,
ookeanide põhjasetted, karbonaatsed kivimid, jne) aga need pole ringes.
Näeme, et olulised vood on süsiniku liikumine taimedesse fotosünteesil (120 ühikut aastas
maismaal ja 107 ühikut aastas ookeanis) ning süsiniku vabanemine süsihappegaasina
atmosfääri hingamise käigus. Taimne hingamine tagastab maismaal sealjuures atmosfääri
umbes poole fotosünteesil fikseeritud süsinikust, ülejäänud naaseb atmosfääri läbi
mullahingamise (laguahelas).
75
2. VEERINGE
Uurige iseseisval allpool olevat skeemi ja olge valmis seda kommenteerima. Vaadake, kuidas
on evaporatsioon ja sademed jaotunud maismaa ja ookeani vahel. Pange tähele, millised on
peamiised veeringe fondid
3. FOSFORIRINGE
Fosforiringe on selle poolest omapärane, et siin puudub täiesti atmosfääri fond. Kogu
kättesaadav fosfor on pärit maakoorest.
76
4. LÄMMASTIKURINGE
Lämmastiku oluline fond on atmosfäär. Sealt satub lämmastik orgaanilisse ainesse peamiselt
tänu molekulaarset lämmastikku fikseerivatele mikroobidele. Osad neist elavad sümbiondina
taimede juurtes (näiteks mügarbakterid perekonnast Rhizobium liblikõielistes taimedes)
kooselus seensümbiondiga (tsüanobakterid samblike koosseisus), osad elavad vabalt (eelkõige
tsüanobakterid ehk sinivetikad, samuti bakterid perekonnast Azotobacter).
Kuna lämmastikku sisaldavad ioonid (nitritioon, nitraatioon, ammooniumioon) on reeglina
hästi dissotseeruvad, on lämmastik väga liikuv element. Organismides on lõviosa lämmastikust
valkude koosseisus (meenutagem, et igas aminorühmas on lämmastikuaatom). Laguahelas
hoolitsevad immobiliseeritud lämmastiku ringesse toomise eest esimesena ammonifitseerijad
bakterid. Ammonifikatsioon on aminorühma (-NH2) sisaldavate orgaaniliste ainete
bakteriaalne lagundamine (aminohapete desamiinimine). Ammonifikatsiooni lõpp-produkt on
ammoniaak (NH3, vesilahuses esineb ammooniumioonina – NH4
+). Aeroobses keskkonnas
ammoniaak ei kogune vaid assimileeritakse koheselt nitrifitseerijate bakterite poolt.
Nitrifikatsioon on ammooniumiooni kaheastmeline oksüdeerimine, algul nitritiooniks (NH4
+
® NO2-, seda teevad näiteks bakterid perekonnast Nitrosomonas), pärast nitraatiooniks
(NO2
- ® NO3-, seda toimetavad näiteks perekonna Nitrobacter esindajad).
Nii ammooniumioonid kui nitrit- ja nitraatioonid on taimedele kättesaadavad.
Anaeroobsetes (e. anoksilistes) tingimustes, mida peamiselt kohtab maismaal liigniisketes
tingimustes ja veekogudes, leiab aset protsess, mille käigus orgaanilisest ainest pärit lämmastik
uuesti atmosfäärifondi naaseb – denitrifikatsioon. Sellega tegeleavad denitrifitseerijad
bakterid, näiteks Pseudomonas denitrificans. Denitrifikatsioon on mineraalne hingamine,
milles orgaanilise aine vesinikult võetavate elektronide lõppaktseptor on nitraat (NO3
-), mis
järk-järgult redutseeritakse kas dilämmastikoksiidiks (N2O) või molekulaarseks lämmastikuks
(N2).
77
ÖKOLOOGIA (LTOM .01.105) KORDAMISKÜSIMUSED, kevad 2019. a.
1. Ökoloogia – aine, alajaotused;
2. Ökoloogia põhimõisted – isend (genet, kloon, ramet), populatsioon, kooslus, ökosüsteem,
bioom;
3. Ökoloogilised tegurid (nende erinevad liigitused), ökoloogiline amplituud,
tolerantsuskõver, ökoloogiline nišš;
4. Ökoloogiline nišš, selle graafiline kujutamine, n
-mõõtmeline eukleidiline niširuum, selle
teljed ehk niši dimensioonid;
5. Liebigi seadus;
6. Ressursid: radiatsioon (PAR, selle omadused ja lainepikkused), CO2, mineraalsed
toitained, vesi,
hapnik, C3 ja C4 taimed, CAM fotosüntees sukulentidel;
7. Tingimusfaktorid: temperatuur, kliima, pH, soolsus, bioomide asend sademete hulga ja
aasta keskmise temperatuuri poolt määratud kahemõõtmelises ruumis;
8. Fenotüübilise varieeruvuse komponendid, fenotüübiline plastilisus, näited;
9. Ressursside klassifikatsioon sünergeetilise efekti järgi;
10. Üksiku populatsiooni kasv, seda kirjeldavad võrrandid – eksponentsiaalne e. piiramatu ja
logistiline e. sigmoidne kasvukõver, keskkonna kandevõime (K), biootiline potentsiaal ehk
erikasvukiirus (r);
11. K
- ja r-strateegid, lõivsuhte mõiste ja näited;
12.
Liigisisene
konkurents,
sümmeetriline
ja
asümmeetriline
konkurents;
13. Konstantse saagi seadus ja
-3/2 astme ehk isehõrenemise seadus, nimetatud seaduste
põhieeldused;
14. Populatsiooni iseloomustavad parameetrid
- populatsioonitihedus, puhas kasvukiirus,
elumus, suremus, viljakus, paljunemisväärtus, Deevey kõverad, demograafilised püramiidid;
15. Populatsioonide levik (isendite jaotus ruumis, viimase tüübid, klonaalsus, sotsiaalne kari, «
isekas kari »), levimine, migratsioon (üheotsapilet, edasi
-tagasi pilet, korduvkasutusega pilet,
näited);
16. Populatsioonidevaheliste interaktsioonide liigitus, näited;
17. Konkurents – mõiste, liigid, Gause eksperimendid, Gause reegel, limiteeriva sarnasuse
printsiip, niššide diferentseerumine;
18. Mutualismi liigid, sümbiontse mutualismi liigid, molekulaarse lämmastiku fikseerijad,
mükoriisa, selle tüübid, näited
19. Kisklus, herbivooria. Kiskja
-saakloom dünaamika Lotka-Volterra tüüpi võrrandsüsteemide
kohaselt;
20. Saakloomade kaitsekohastumused, mimikri tüübid;
21. Parasitism, parasiitide liigitused, parasitoidid, mikroparasiitide puhast kasvukiirust
mõjutavad tegurid, parasiidi levikulävi;
22. Kommensalism, laguahel, lagundajad, nende klassifikatsioon suuruse järgi; mikro
- meso-
ja makrofauna suhteline osatähtsus laguahelas eri kliimavööndites;
23. Koosluste struktuuri aspektid, vertikaalne ja horisontaalne struktuur, diversiteet;
24. Organitsistlik (Clements) versus individualistlik (Gleason) paradigma sünökoloogias,
koosluste ordinatsioon ja klassifikatsioon;
78
25. Eesti metsakasvukohatüüpide klassifikatsiooni skeem;26. Koosluste primaarne ja
sekundaarne suktsessioon, soode suktsessioon ja tüübid, kliimaksi mõiste; 27. Bioloogilise
mitmekesisuse mõiste, taksonoomiline mitmekesisus;
28. Liikide ligikaudne arv maakeral ja tähtsamates, taimekoosluste liigirikkuse
maailmarekordid; 29. Liikide arvu varieerumine eri organismirühmades geograafilise laiuse,
absoluutse kõrguse ja sügavuse gradiendil;
30. Lokaalsed geograafilised diversiteedigradiendid – poolsaare efekt, lahe efekt küürselg
-
kõver (hump
-backed curve;
31. Liikide arvu ja pindala seos;
32. Tähtsamate organismirühmade liikide (ja kõrgemate taksonite) arvu muutused
evolutsioonilises ajas. Liikide väljasuremise dünaamika, massekstinktsioonid;33. Haeckel’i
seletus troopika suure liigirikkuse kohta, mittetasakaaluline käsitlus (+ häiringute roll
mitmekesisuse säilimisel);
34. Primaarproduktsiooni globaalne jaotus maismaal ja meres;
35. Energiavoog ökosüsteemides. Troofilised tasemed, ökoloogilised püramiidid, toiduahel,
toiduvõrgustik;
36. Ttroofiliste tasemete vaheline energia ülekande efektiivsus;
37. Erinevate troofiliste tasemete ja erinevate organismirühmade produktsiooniefektiivsus ja
assimilatsiooniefektiivsus;
38. Aineringed – süsinikuringe, veeringe, lämmastikuringe, fosforiringe. Põhilised fondid ja
vood ringetes;
39. Miks ma pole piisava põhjalikkusega eksamiks valmistunud?
Ökoloogia kursuse põhjalik konspekt. Konspekteeritud on kõik 14 loengut.
Sarnased õppematerjalid
![ÖKOLOOGIA eksami küsimuste vastused]()
13
doc
ÖKOLOOGIA eksami küsimuste vastused
ÖKOLOOGIA (LOOM .01.105) KORDAMISKÜSIMUSED, kevad 2011. a.
1. Ökoloogia aine, alajaotused;
Teadus, mis käsitleb organismide ja keskkonna suhet. Kõikide sidemed kõikidega.
Jaguneb:
a) Ökofüsioloogia e molekulaarne ökoloogia
b) Autökoloogia (isendi/organismi tasandil)
c) Demökoloogia (populatsiooni tasandil)
d) Sünökoloogia (eluskoosluse, populatsioonide tasandil)
e) Süsteemökoloogia (ökosüsteemi tasandil, elus kooslus + eluta keskkond)
f) Biosfäroloogia e biosfääri ökoloogia (globaalne ökosüsteem)
2. Ökoloogia põhimõisted isend (genet, kloon, ramet), populatsioon, kooslus, ökosüsteem,
bioom;
Isend- kindla genotüübiga organism
![Ökoloogia konspekt]()
13
doc
Ökoloogia konspekt
1. Aine, alajaotused (allpool) , areng.
Ökoloogia - teadus, mis uurib elusa ja eluta looduse omavahelist suhet, ei keskendu ühele
objektile, vaatleb tervikut.
E. Haeckel 1869 ökoloogia on teadus organismide ja kk suhetest.
E. Odum teadus looduse struktuurist ja funktsoonist.
2. Ökoloogia põhimõisted.
Ökoloogia valdkonnad:
1) Organelli tase
2) Raku tase (ainurakse puhul isend)
3) Koe tase
4) Organi tase
5) Isendi tase autökoloogia, uurib abiootilisi kk faktoreid.
6) Populatsiooni tase demökoloogia e. populatsiooni ökoloogia.
7) Koosluse tase kooslusökoloogia e. sünökoloogia, uurib mitmeliigilisi pop. süsteeme.
![Ökoloogia kordamisküsimused 2022]()
31
docx
Ökoloogia kordamisküsimused 2022
Panmiktiline populatsioon ehk deem – vabalt ristuvate isendite kogum, pole
populatsioonisiseseid paljunemisbarjääre.
Levis ehk diaspoor – on igasugune organismide (eriti taimede ja seente) levimisalge, ka
organismi osa, mille abil organismid levivad (eosed ehk spoorid, seemned).
�Bioom ehk makroökosüsteem – sarnastes klimaatilistes ja ökoloogilistes tingimustes levivad
ning sarnase struktuuriga ökosüsteemid üle maailma.
Ökofüsioloogia – ökoloogia piirteadus, mis uurib ökoloogilisi protsesse organelli ja organi
tasemel.
Autökoloogia – ökoloogia valdkond, mis tegeleb uuringutega isendi tasemel.
Demökoloogia – ökoloogia valdkond, mis tegeleb uuringutega populatsiooni tasemel.
Sünökoloogia – ökoloogia valdkond, mis tegeleb uuringutega koosluse tasemel.
Süsteemökoloogia – ökoloogia valdkond, mis tegeleb uuringutega kogu ökosüsteemi
tasemel.
� 2. AUTÖKOLOOGIA
![Nimetu]()
42
docx
Nimetu
72. Kuidas avaldub produktsiooniefektiivsus?
A. PE = In/Pa-1
B. PE = An/In
C. PE = Pn/An-1
D. PE = Pn/An
73. Kas kiskja-saaklooma süsteemis:
A. Kiskja populatsioonitiheduse madalseis vastab saaklooma populatsiooni suurimale tihedusele
B. Kiskja ja saaklooma populatsioonitihedus võnguvad samas rütmis
veerandperioodilise faasnihkega
C. Saaklooma populatsioonitihedus on kiskja omast sõltumatu kuid fluktueerib samas rütmis
74. Termini ökoloogia võttis kasutusele:
A. K. Zobel
B. V. Masing
C. Haeckel
D. Hutchinson
E. V. Gause
75. Seen võib olla:
A. Loom
B. Imetaja
C. Nematood
D. Parasiit
E. Primaarne produtsent
76. Produktsiooni efektiivsus on suurem:
A. Kõigusoojastel selgroogsetel?
B. Püsisoojastel selgroogsetel?
C. Nendel selgroogsetel, kelle energiakadu hingamisele on null?
77. Millise elemendi ringe puhul on atmosfäär väheoluliseks fondiks?
A. N
B. P
C. C
D. O
78
![Ökoloogia kordamisküsimused]()
11
doc
Ökoloogia kordamisküsimused
1. Aine, alajaotused (allpool) , areng.
Ökoloogia - teadus, mis uurib elusa ja eluta looduse omavahelist suhet, ei keskejdu ühele objektile,
vaatleb tervikut.
E. Haeckel 1869 - ökoloogia on teadus organismide ja kk suhetest.
E. Odum - teadus looduse struktuurist ja funktsoonist.
2. Ökoloogia pôhimôisted.
Ökoloogia valdkonnad:
1) Organelli tase- uurib olulisi eluavaldusi madalamal str tasemel
2) Raku tase (ainurakse puhul isend)
3) Koe tase
4) Organi tase- autökoloogia, org. Ja keskk. Suhete uurimine isendi tasandil
5) Isendi tase - autökoloogia, uurib abiootilisi kk faktoreid.
6) Populatsiooni tase - demökoloogia e. populatsiooni ökoloogia.
![Ökoloogia konspekt]()
16
doc
Ökoloogia konspekt
Ökosüsteem Esimesena kasutas seda sõna Tansley 1935. eluskooslus ja selle
abiootiline keskkond. Nt kuumaveeallikas, lillepeenar vms.
Bioom sarnaste ökosüsteemide kogum üle maailma. Nt troopiline vihmamets,
tundra.
Biosfäär globaalne ökosüsteem. Kõik teadaolevad ökosüsteemid on avatud
süsteemid, st et nad vahetavad naabersüsteemidega nii ainet kui energiat, ja ei ole
seetõttu eraldiseisvad üksused.
Ökoloogia valdkonnad:
· Molekulaarne ökoloogia meetodile viitav.
· Ökofüsioloogia uurib füsioloogiliste protsesside kohanemist vastavalt
keskkonnale.
· Antökoloogia isendi suhted keskkonnaga.
· Populatsiooniökoloogia, demökoloogia(demograafiline)
· Sünökoloogia(koos, kaasa, kooslemise), kooslusökoloogia
· Süsteemökoloogia
· Geograafiline ökoloogia
· Biosfäroloogia
Ökoloogiline faktor(tegur) aine, energia või informatsiooni voog keskkonnast, mis
![ÖKOLOOGIA kordamisküsimuste vastused 2012]()
18
doc
ÖKOLOOGIA kordamisküsimuste vastused 2012
1. Ökoloogia aine, alajaotused;
Teadus organismide ja nende keskkonna vahelistest seostest (biootiline, abiootiline KK)
1) Molekulaarne ökoloogia (molekul) meetodile viitav.
2) Ökofüsioloogia (molekul, organ, isend) uurib füsioloogiliste protsesside kohanemist vastavalt keskkonnale.
3) Antökoloogia (isend) isendi suhted keskkonnaga.
4) Populatsiooniökoloogia, demökoloogia(demograafiline) (populatsioon)
5) Sünökoloogia, kooslusökoloogia (kooslus)
6) Süsteemökoloogia (ökosüsteem)
7) Geograafiline ökoloogia (bioom)
8) Biosfäroloogia (biosfäär)
2
![ÖKOLOOGIA kordamisküsimused 2013]()
11
doc
ÖKOLOOGIA kordamisküsimused 2013
ÖKOLOOGIA (LOOM .01.105) KORDAMISKÜSIMUSED, kevad 2013. a.
1. Ökoloogia aine, alajaotused;
Ökoloogia on teadus organismide ja keskkonna vahelistest suhetest.
Ökoloogia alajaotused on :
· Ökofüsioloogia (organell, rakk, organ)
· Autökoloogia (isend) organism ja keskkonna suhe isendi tasemel
· Demökoloogia (populatsioon)
· Sünökoloogia (kooslus)
· Süsteemökoloogia (ökosüsteem, biosfäär)
2. Ökoloogia põhimõisted isend (genet, kloon, ramet), populatsioon, kooslus, ökosüsteem, bioom;
· isend organism, mis ei moodusta iseseisvaid mooduleid
o kloon ehk genet geneetiliselt identne moodulite kogum
o ramet taime puhul võsu, risoomiga ühendatud, iseseisva juuresüsteemiga moodul
· populatsioon ühise genofondiga isendite kogum
· kooslus koos eksisteerivad populatsioonid
· ökosüsteem elukooslus ja selle abiootiline kes
Meedia
Kommentaarid (0)



























































































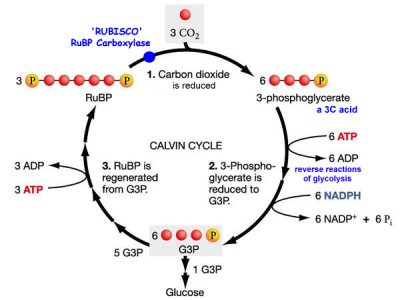

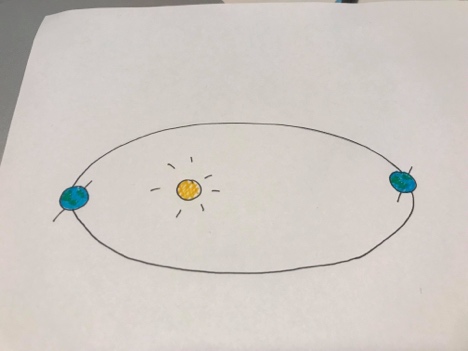
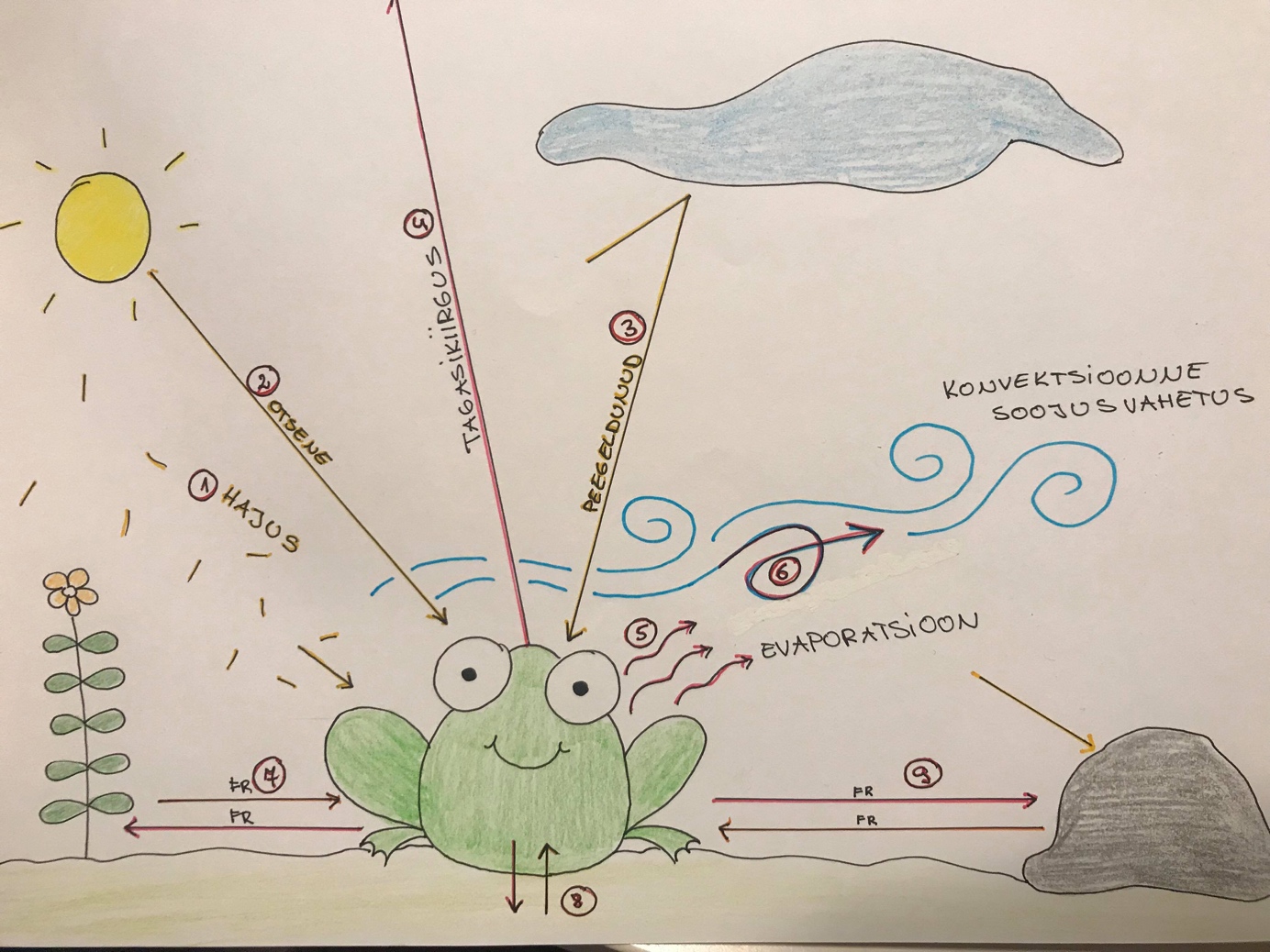
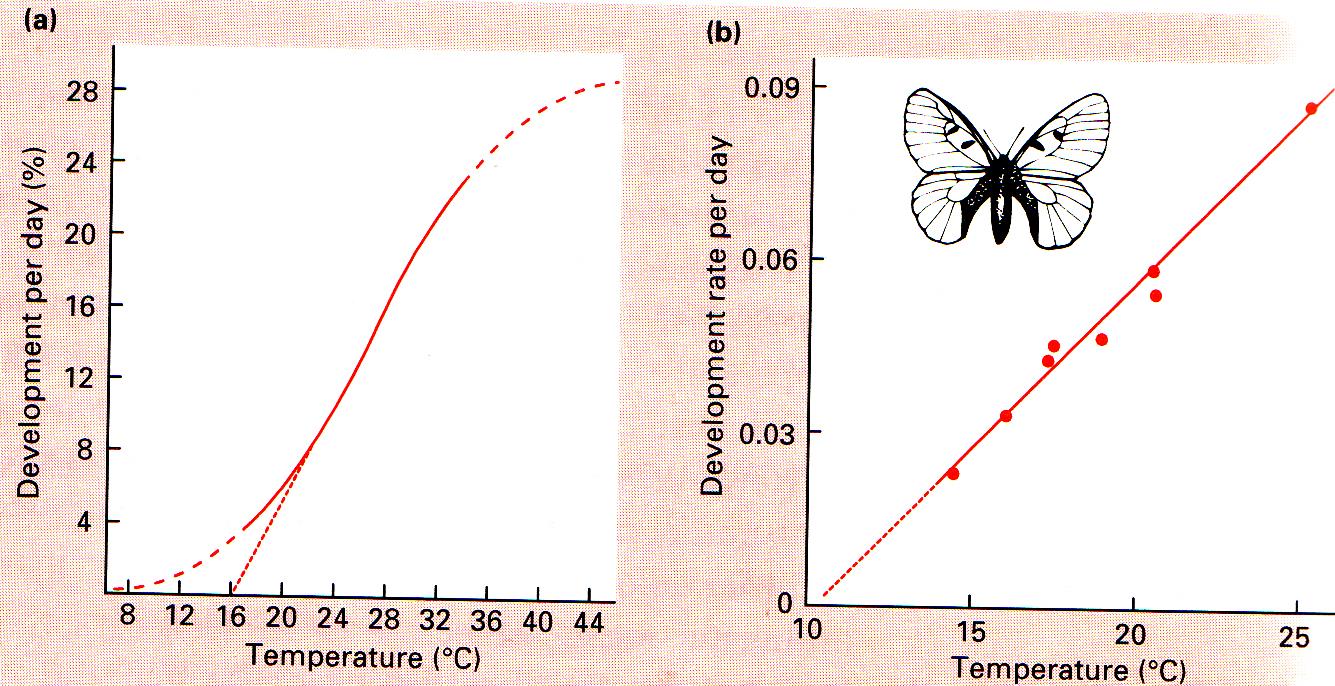
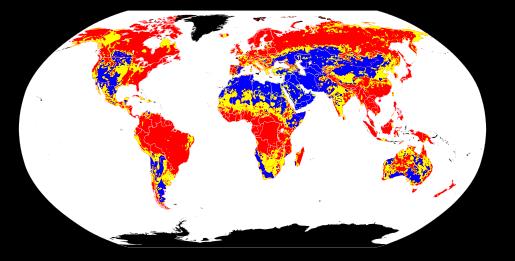
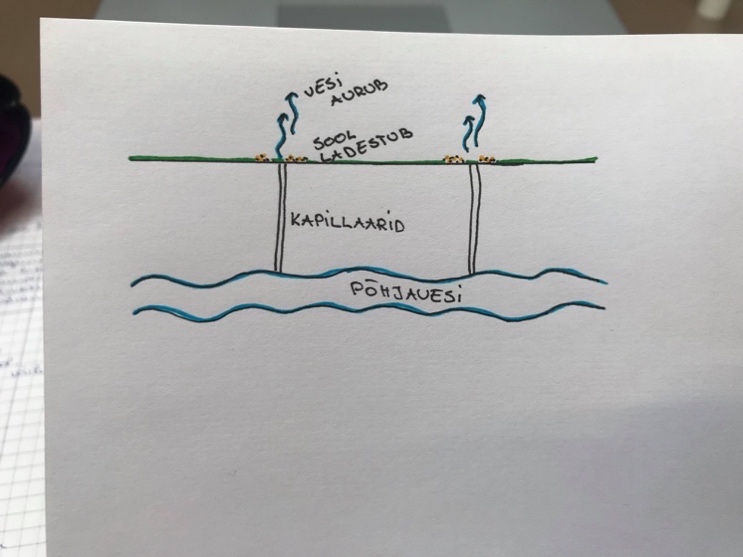
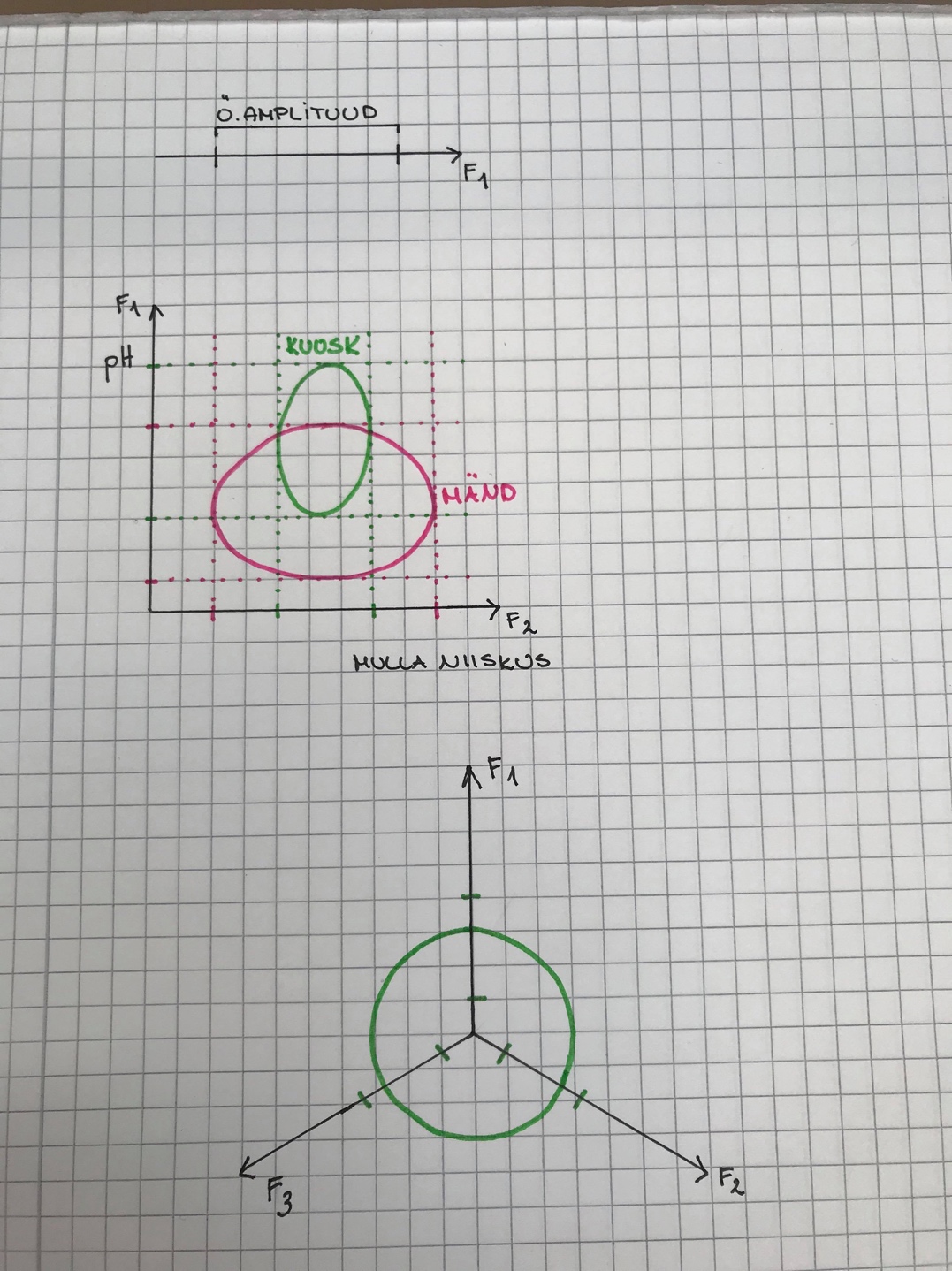
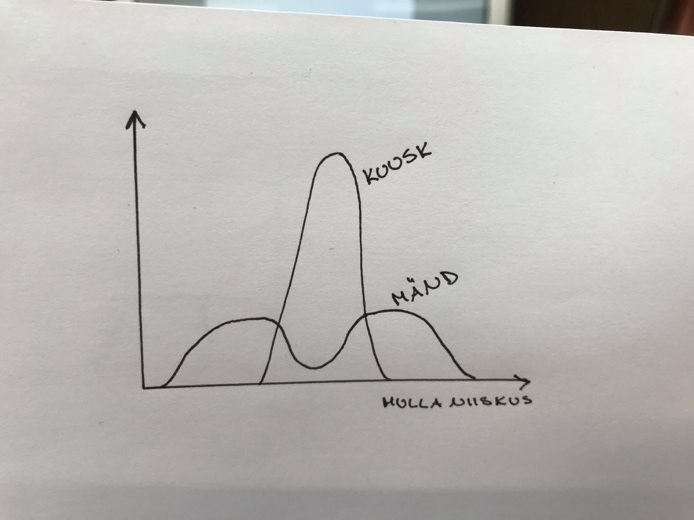
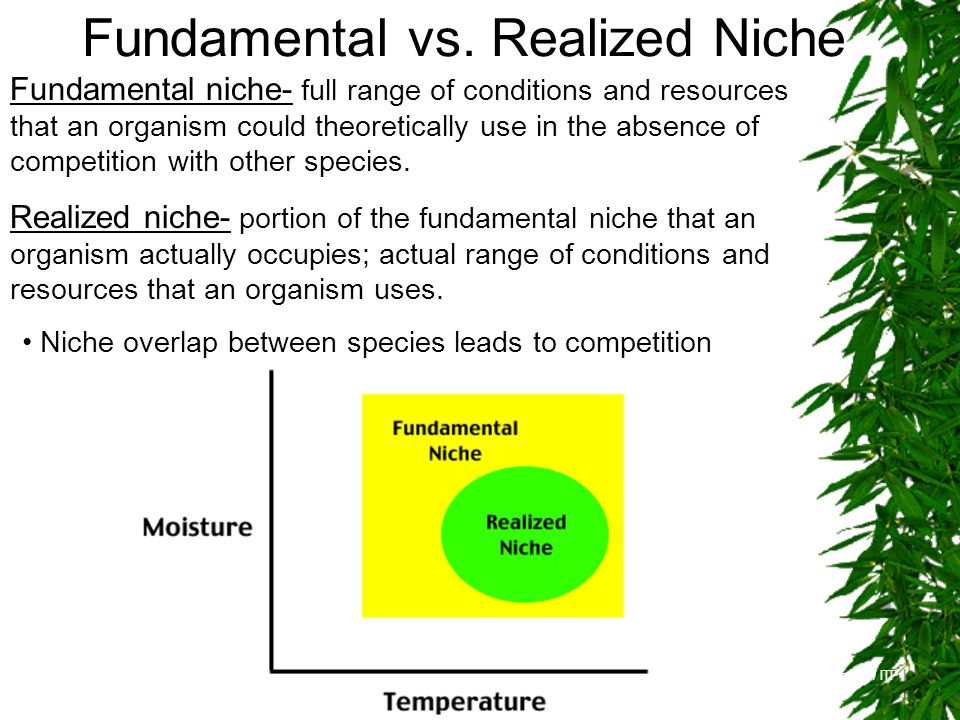
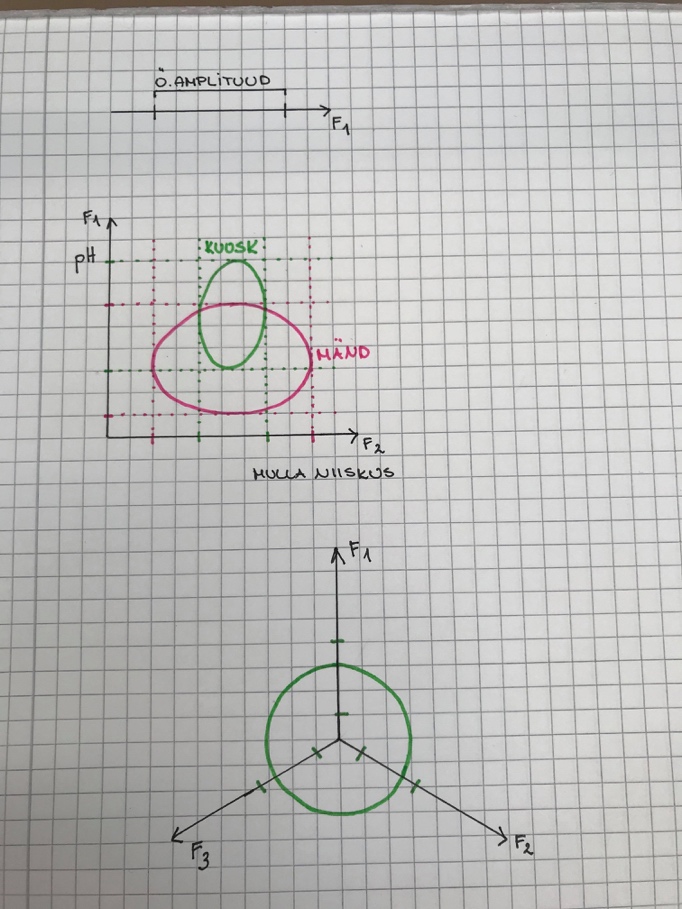
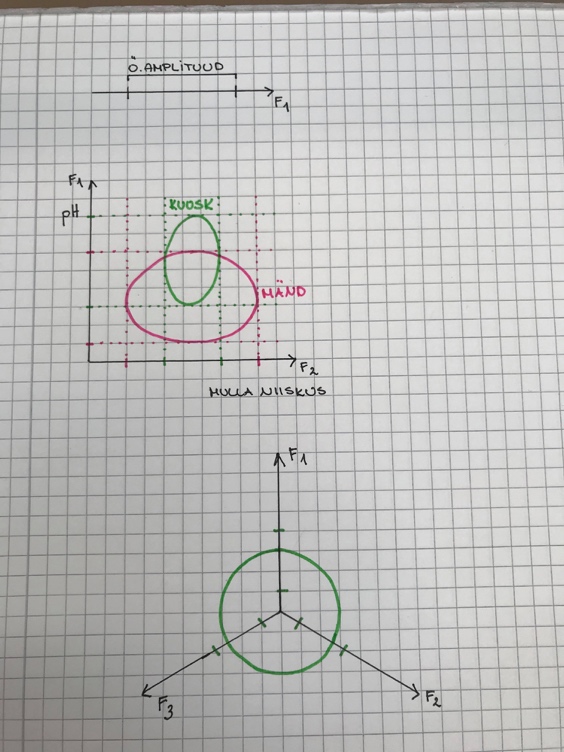
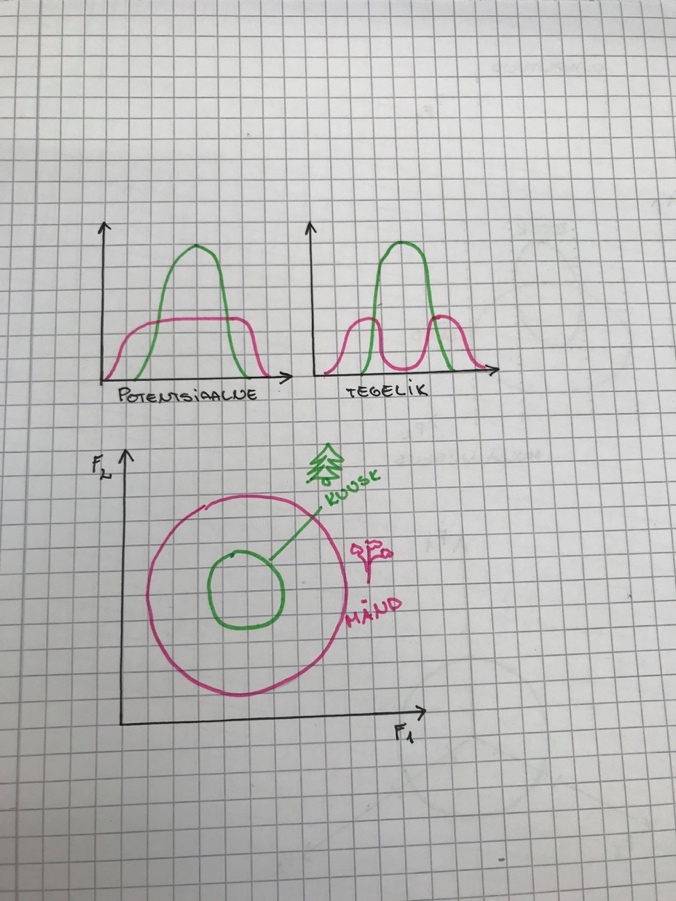



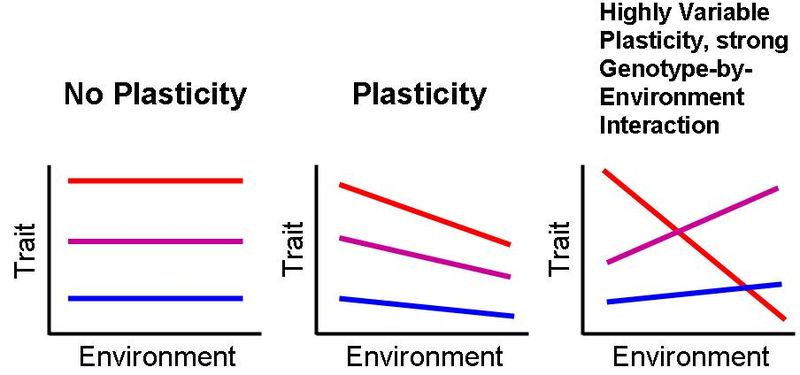
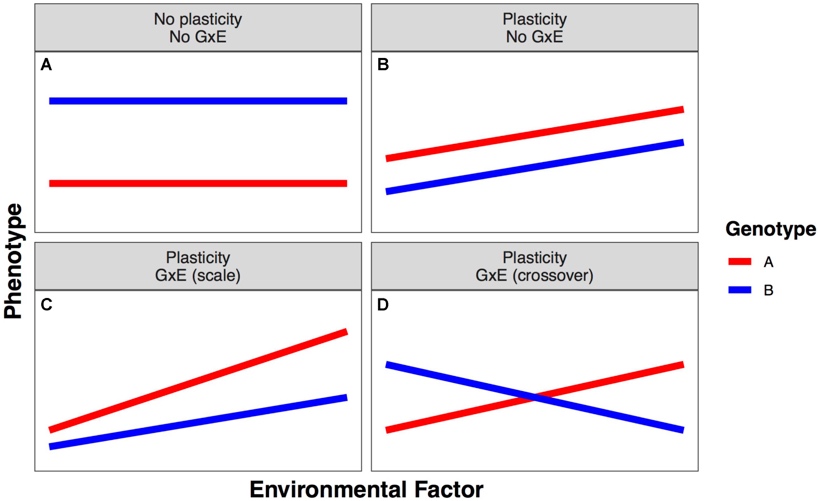
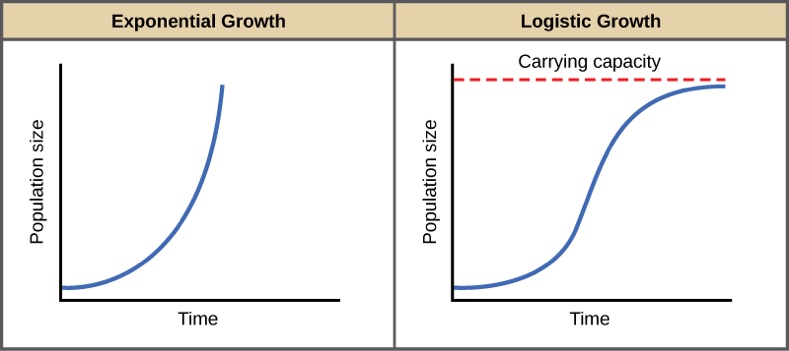
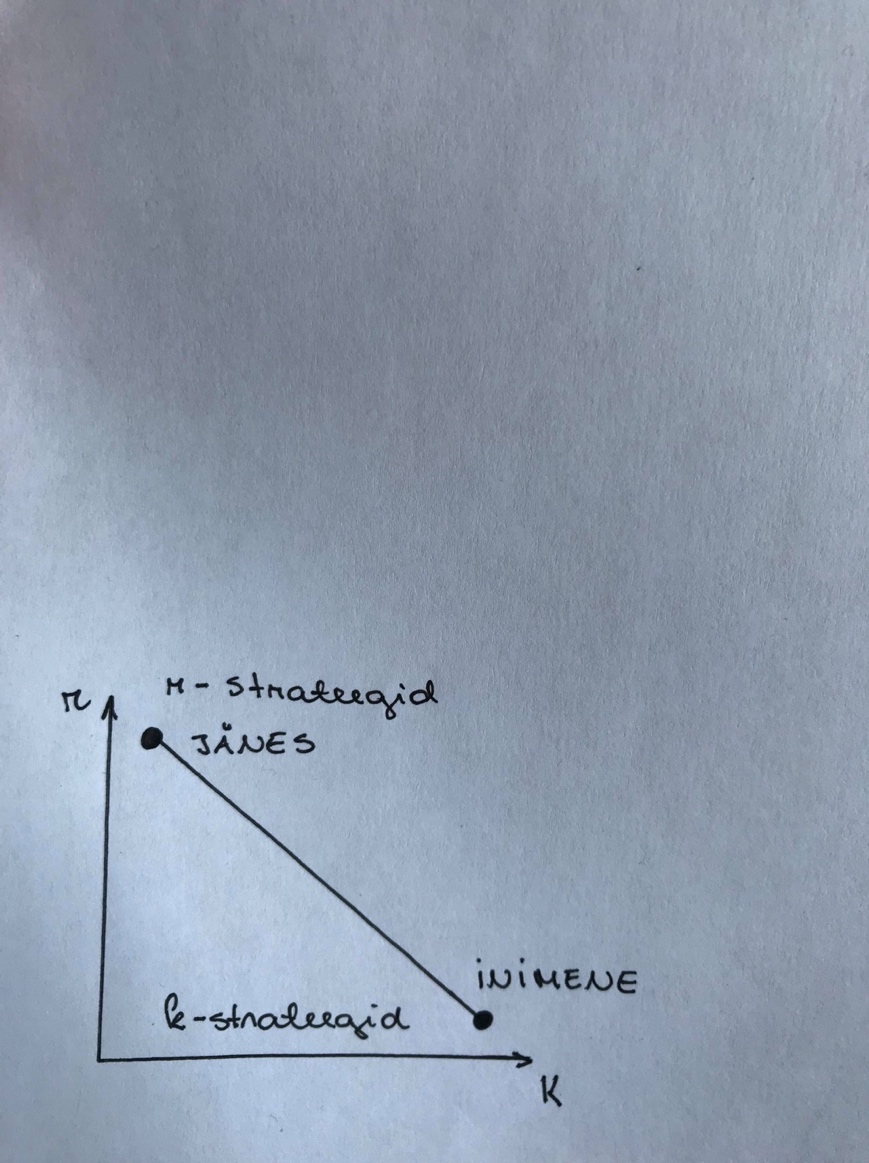
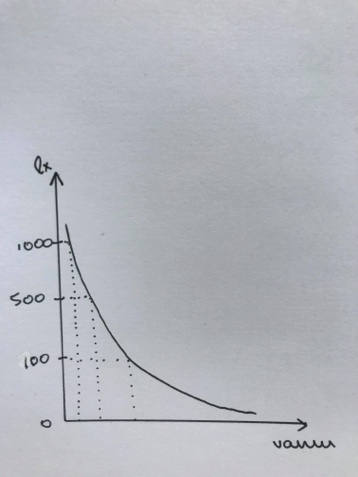

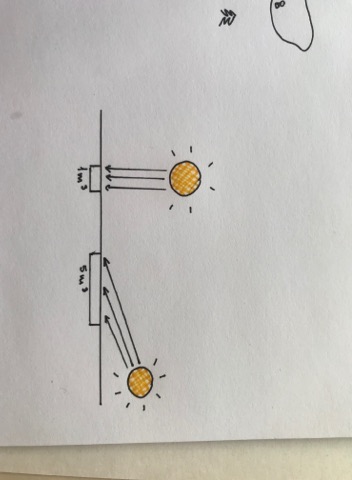
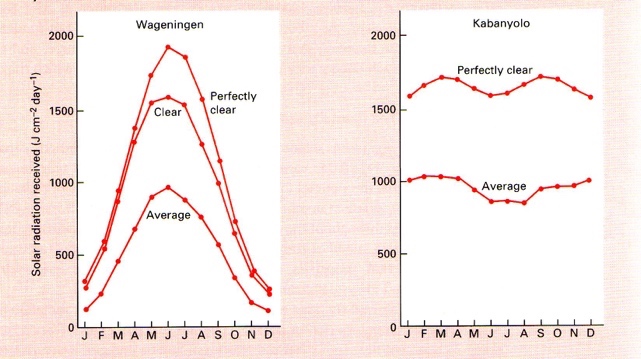
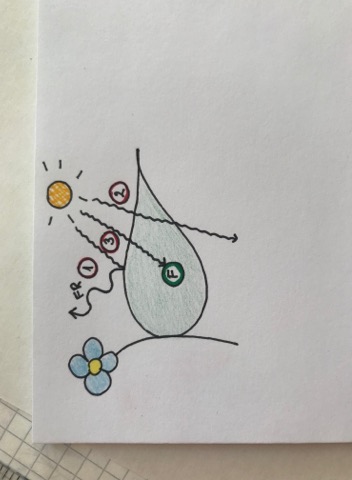
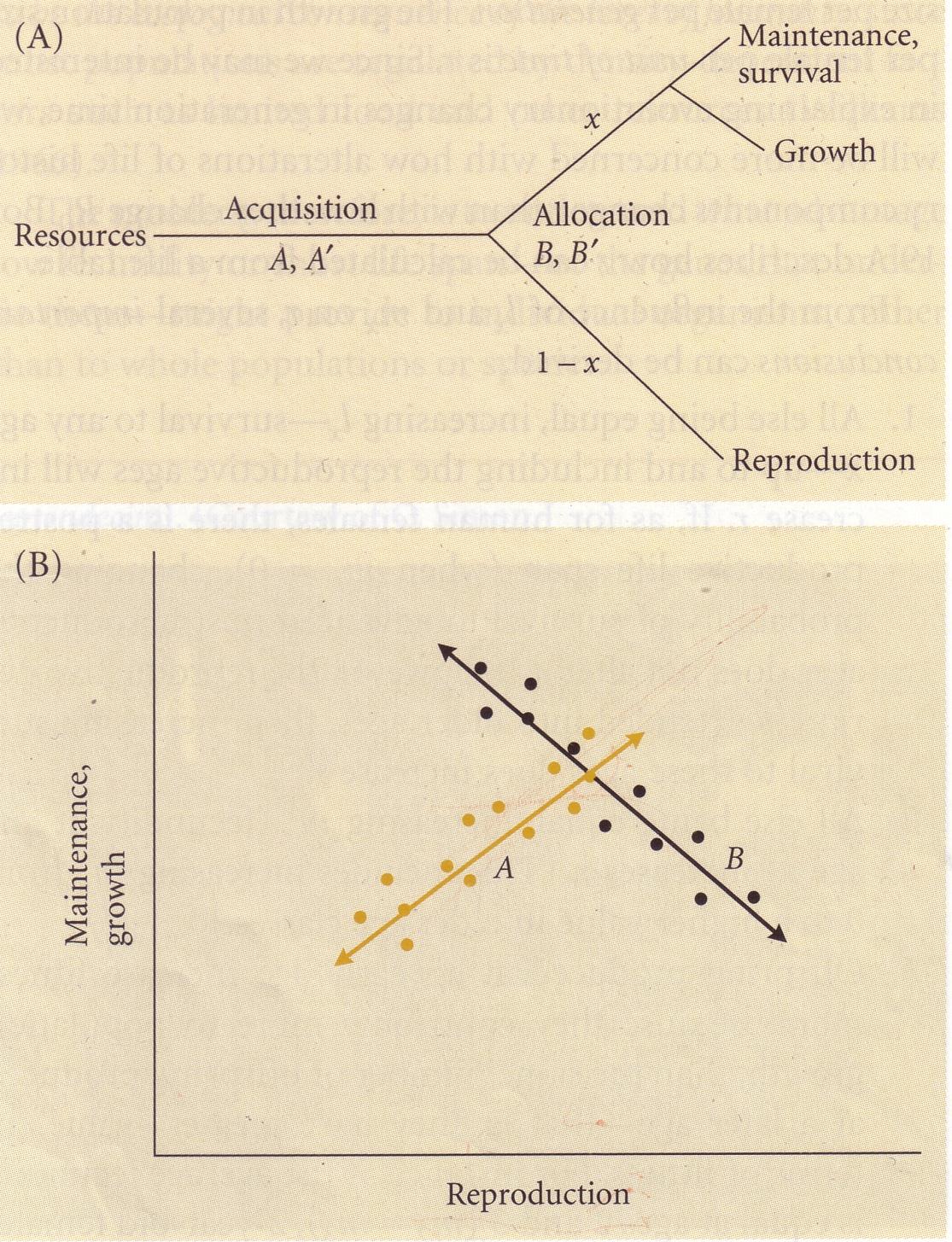
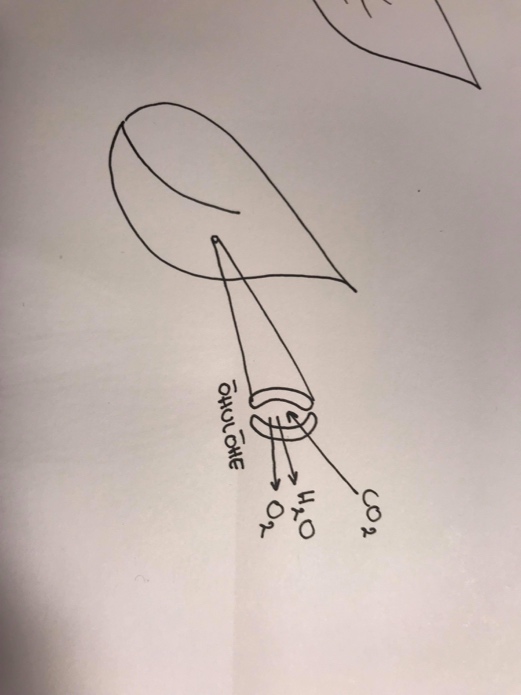
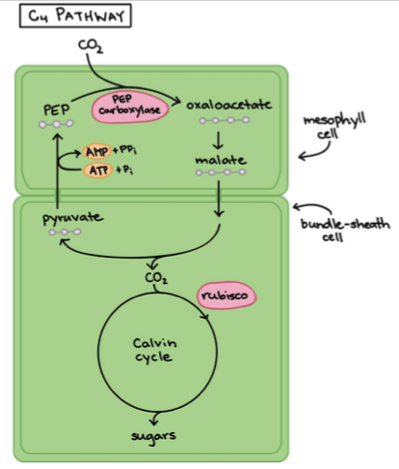
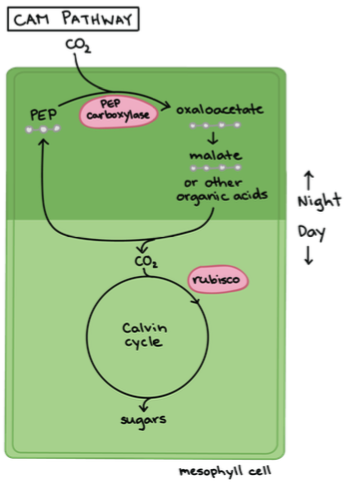
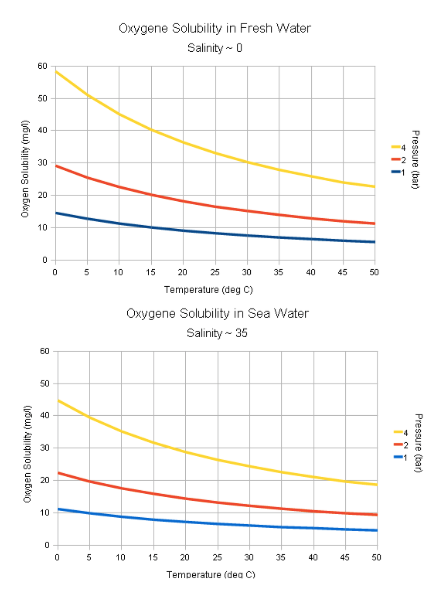
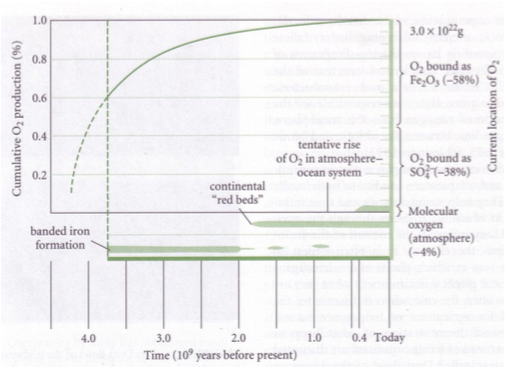
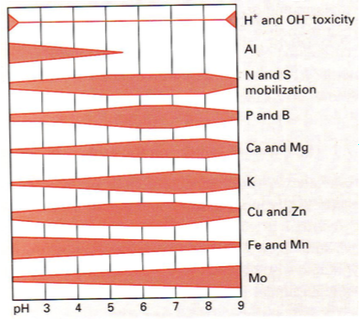
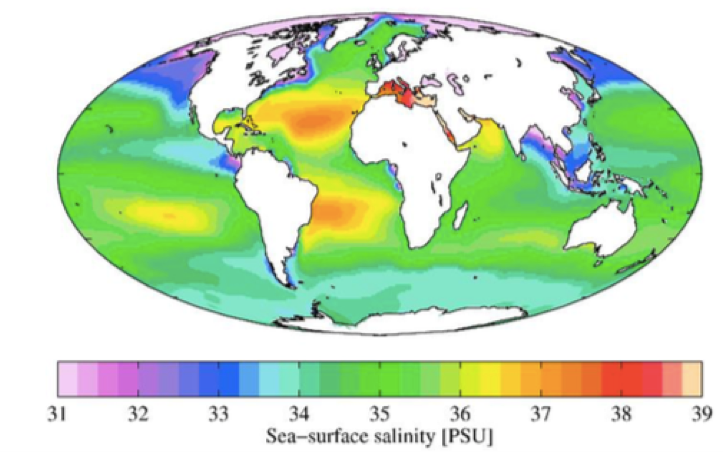
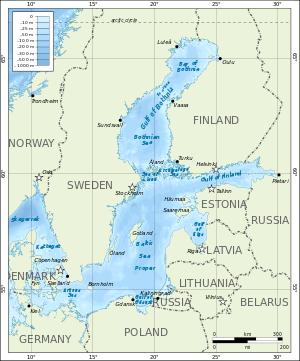
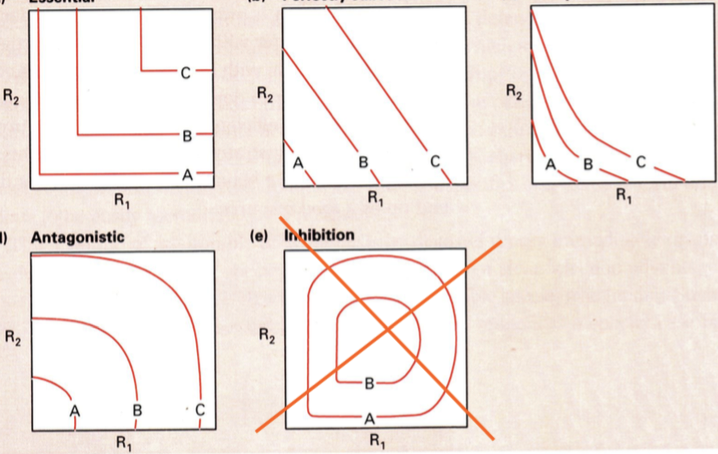
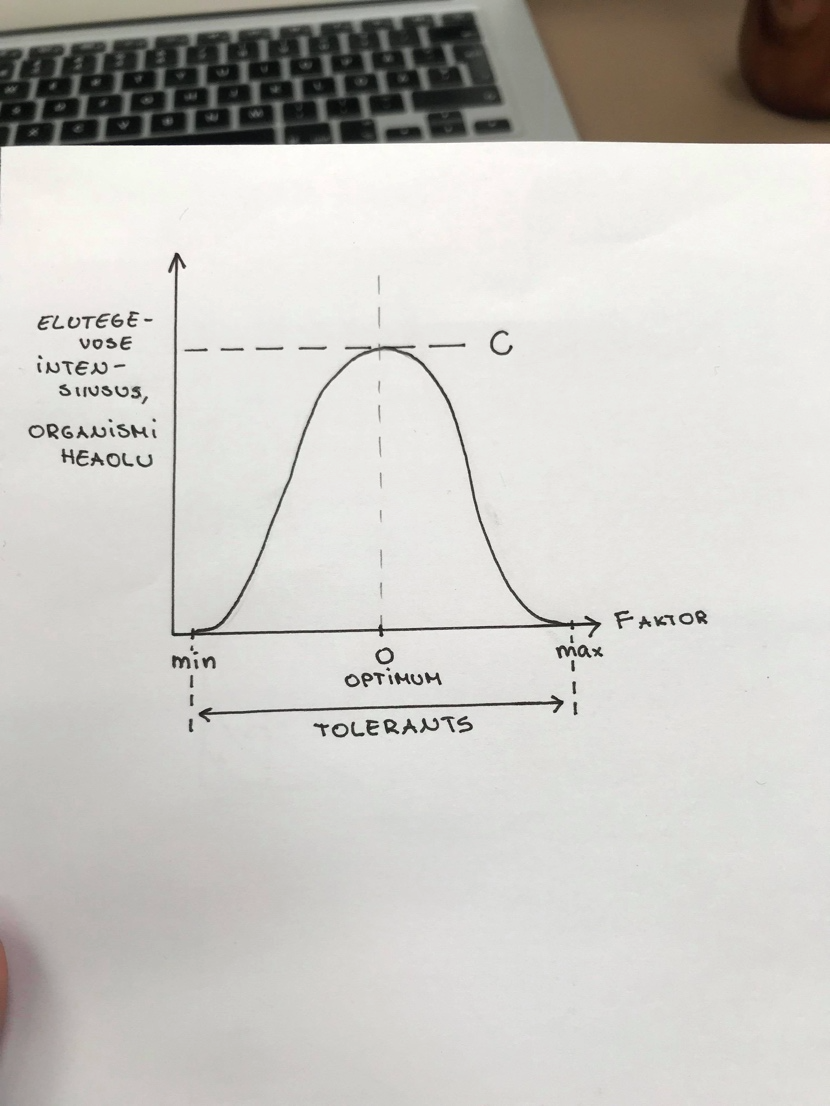

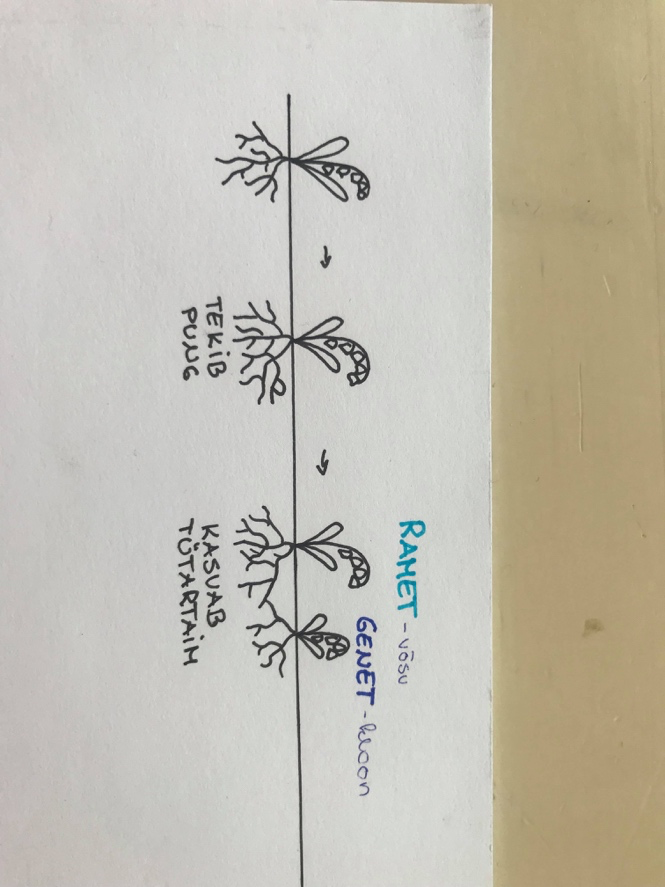
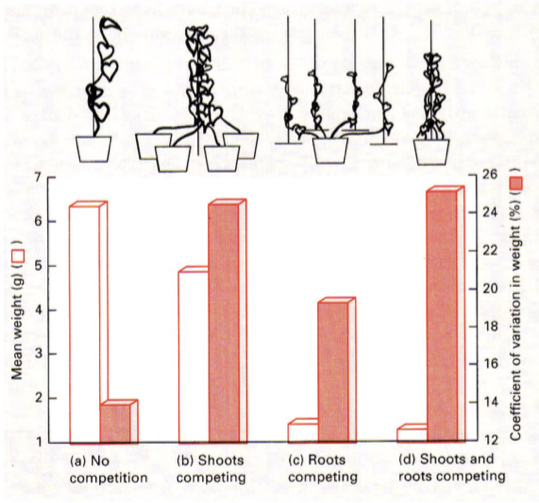
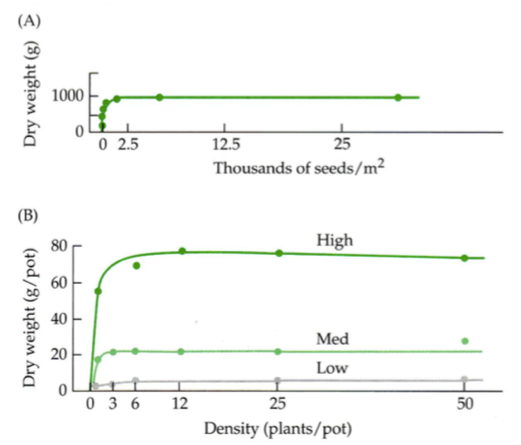
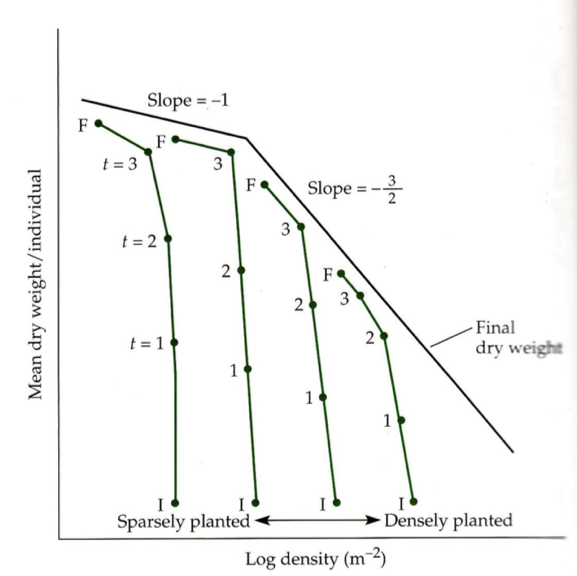
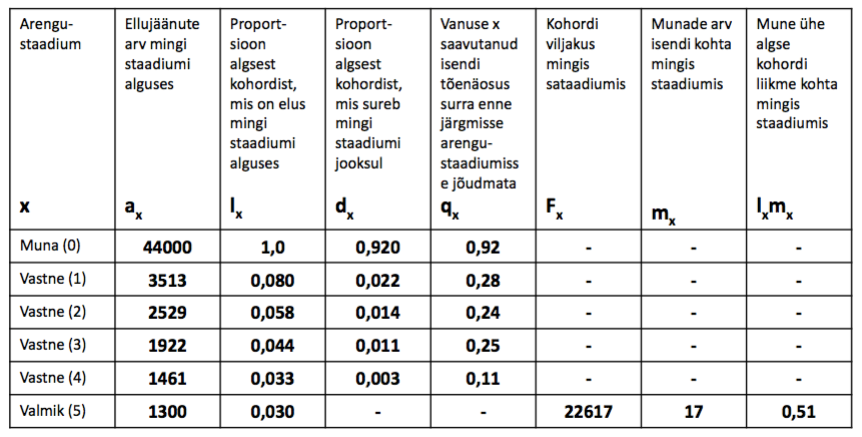
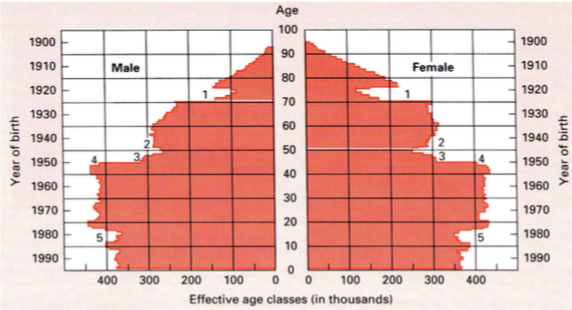
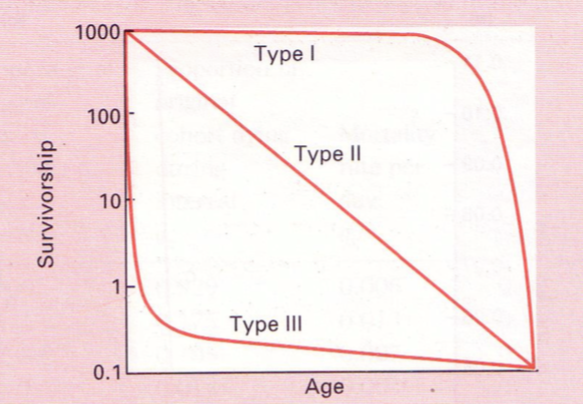
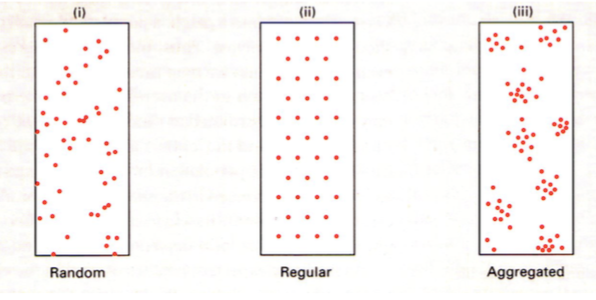
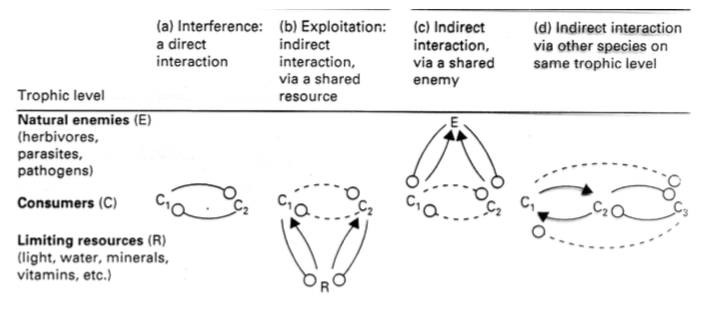
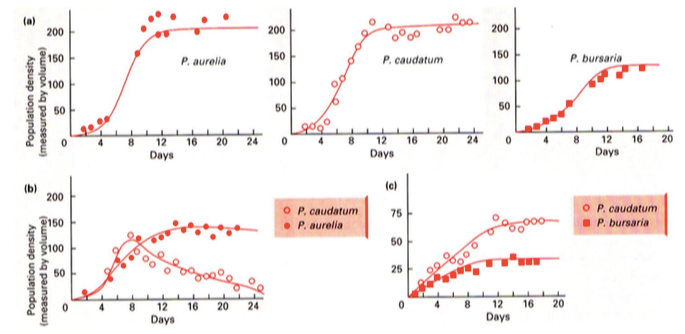
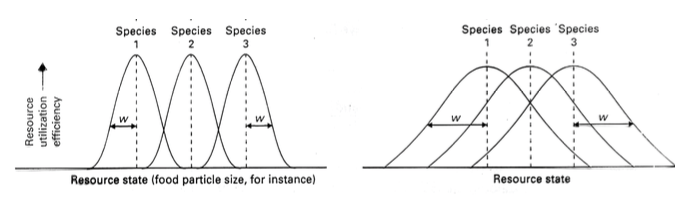
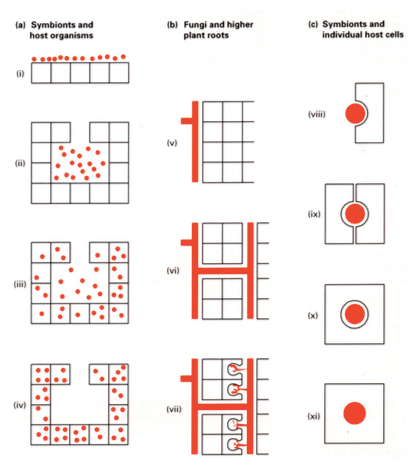
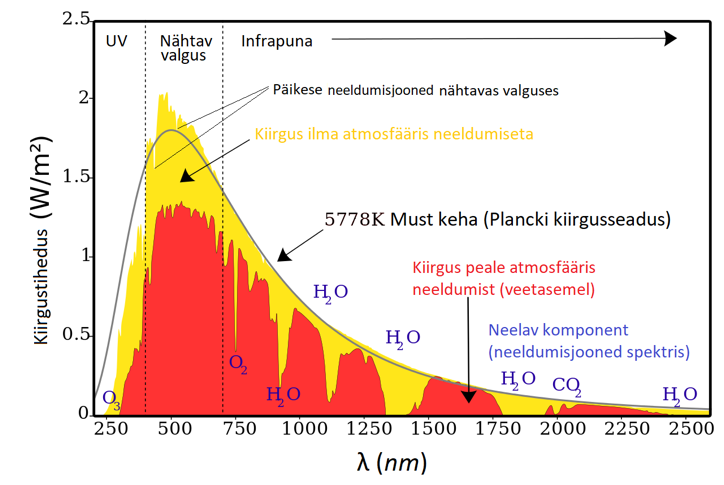



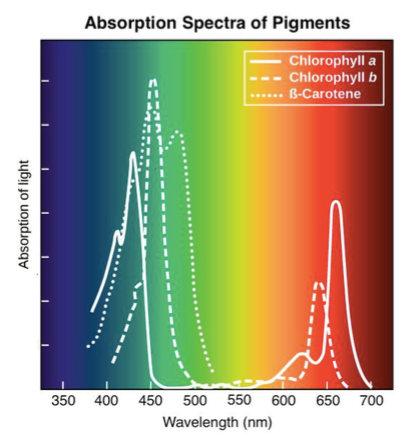
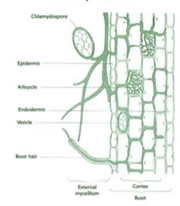

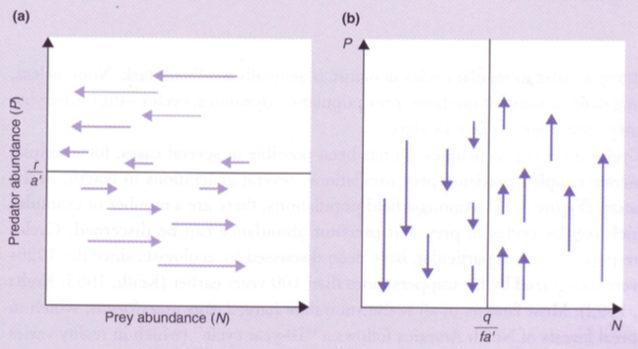
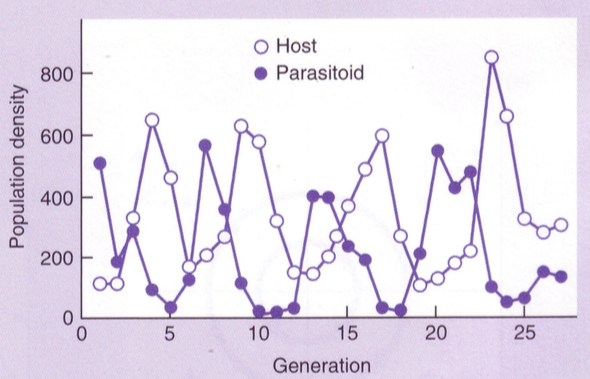
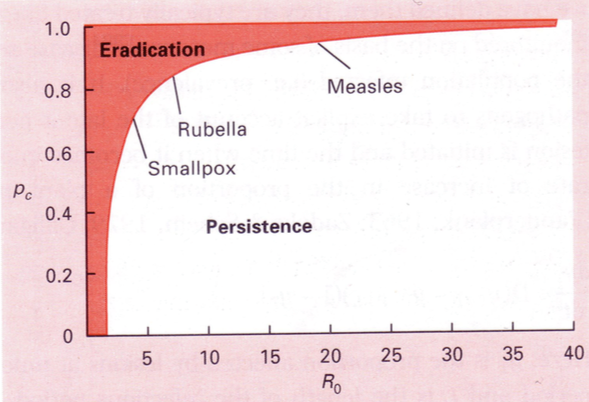
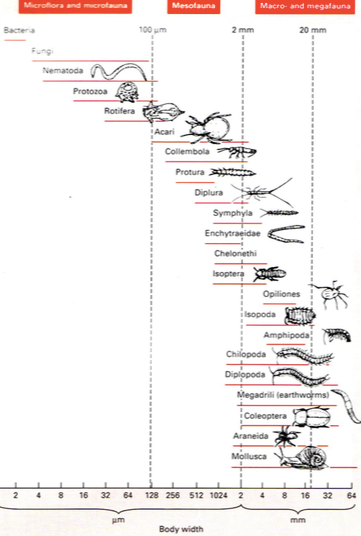
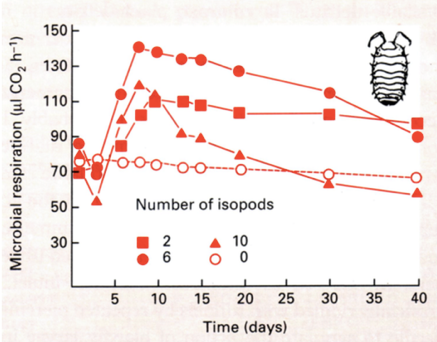
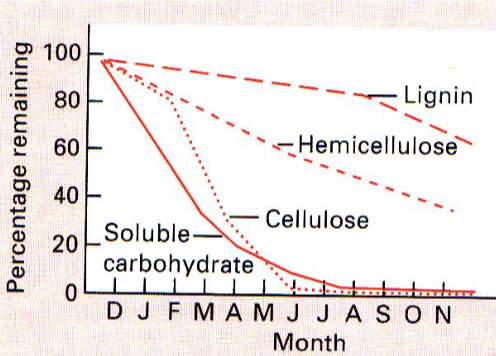
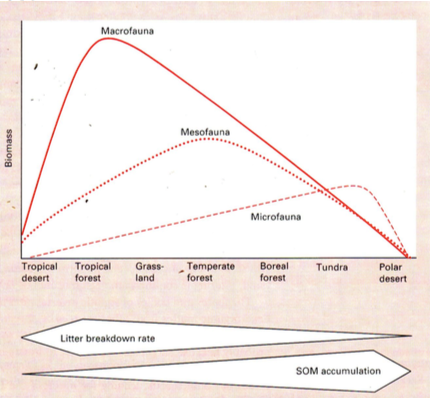
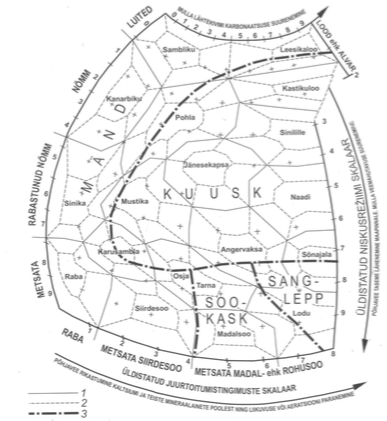
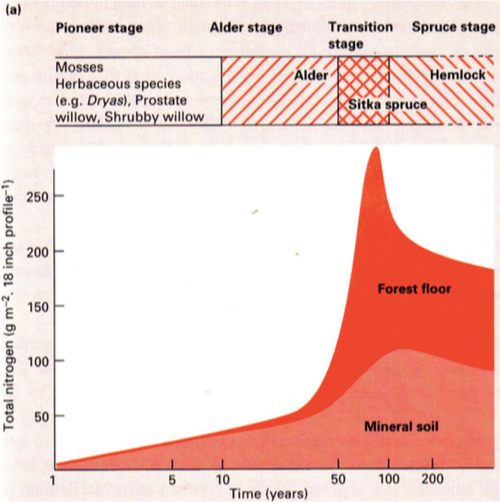
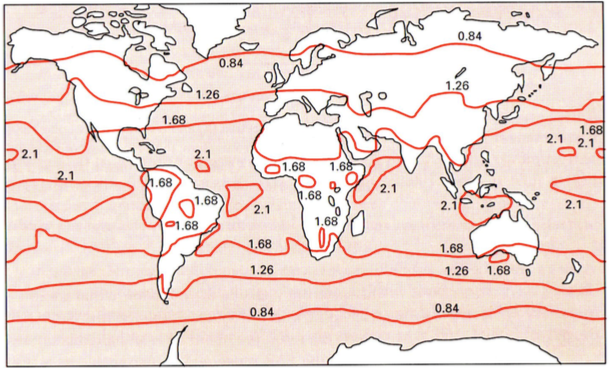
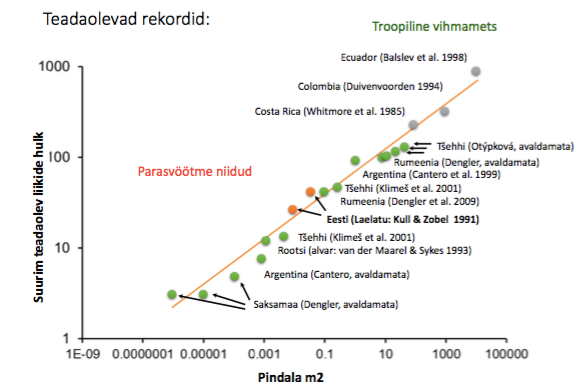
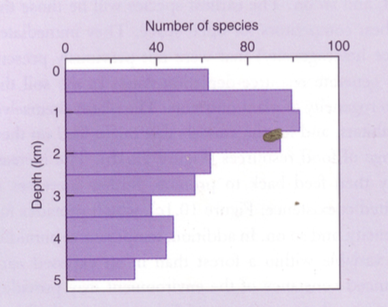
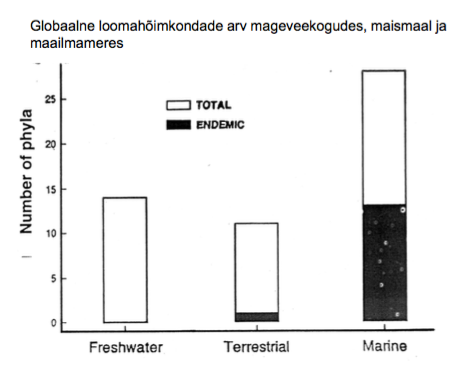
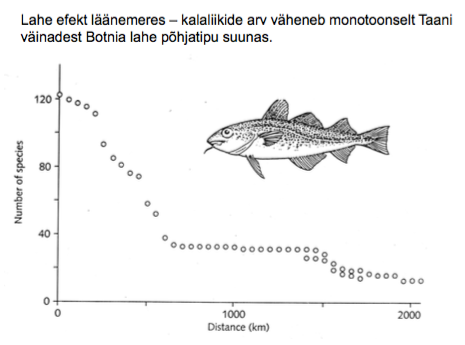
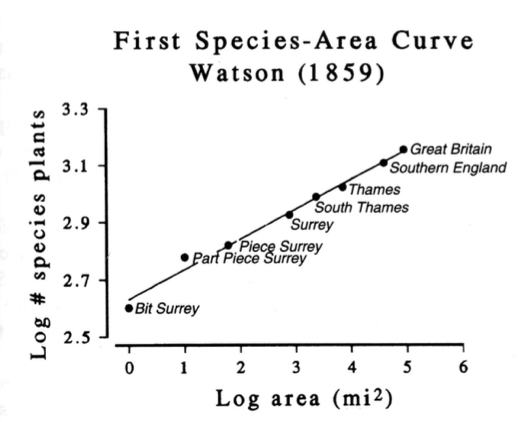
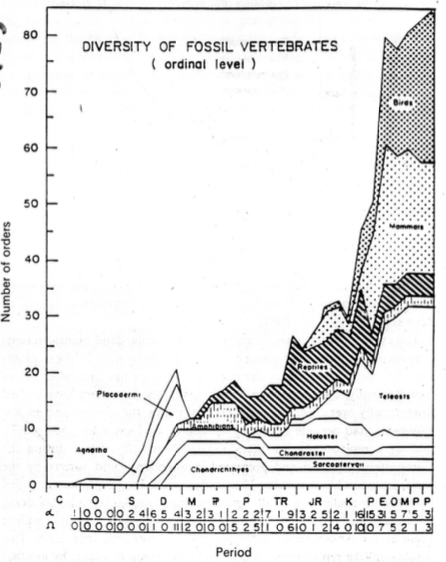
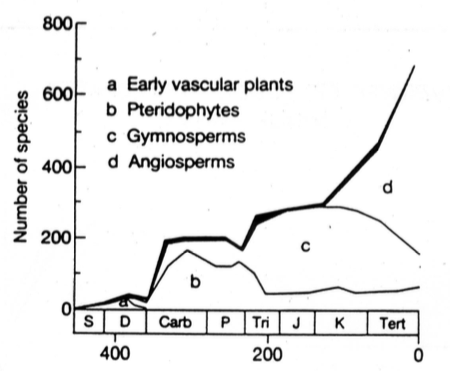
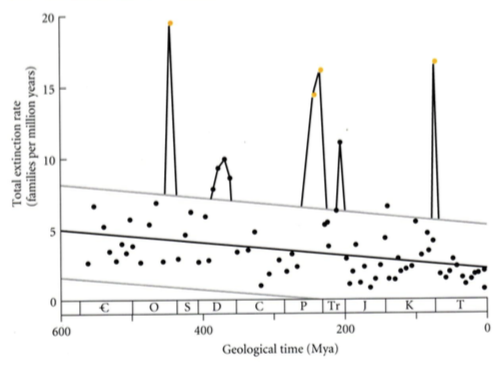
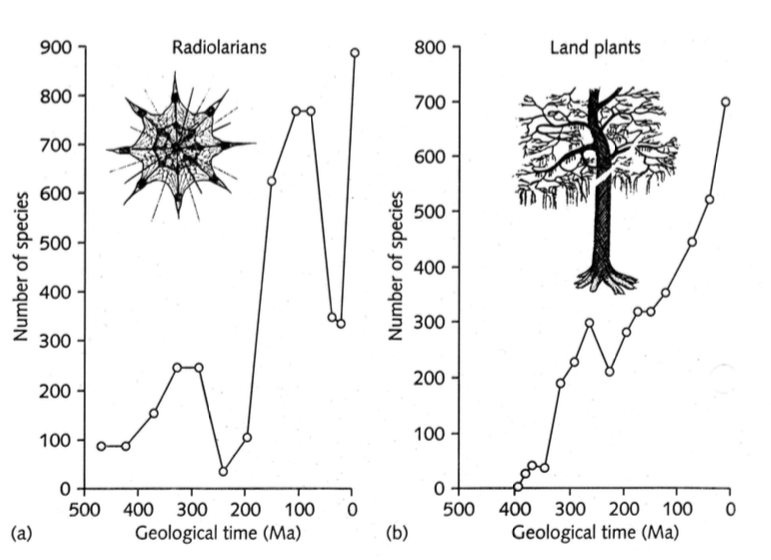
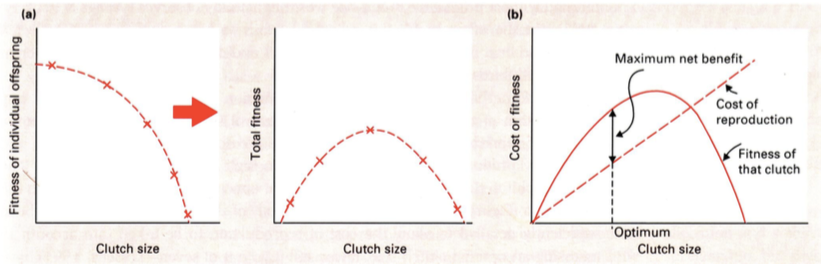
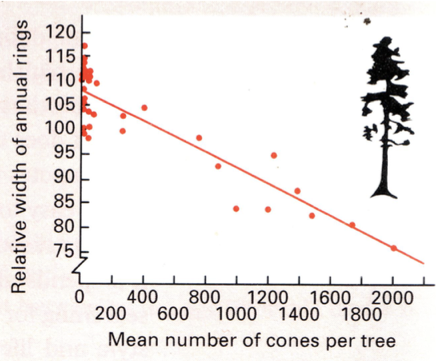
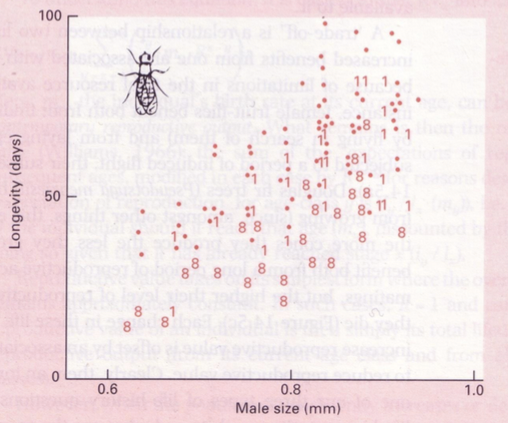
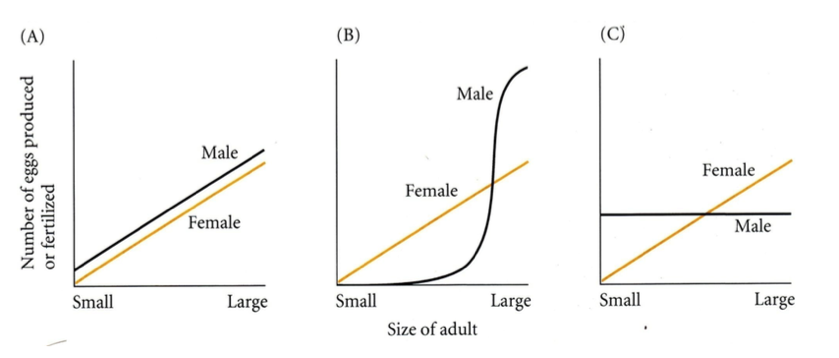
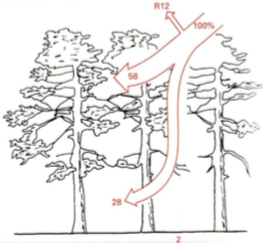
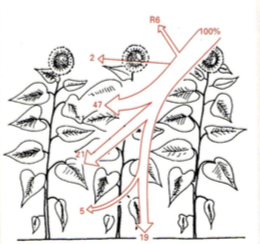
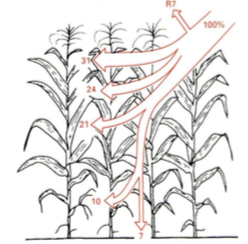
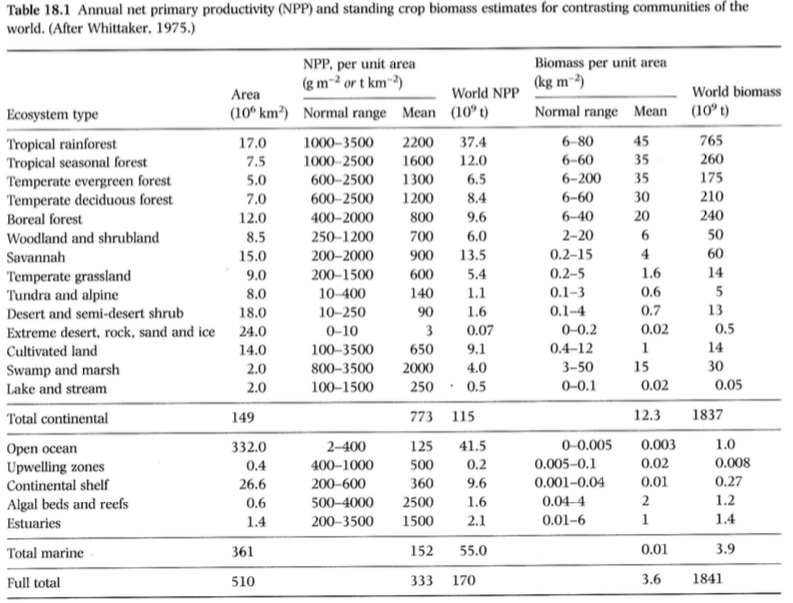
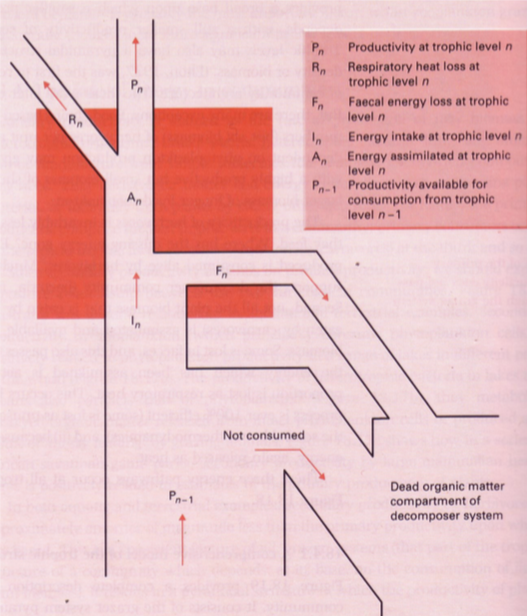
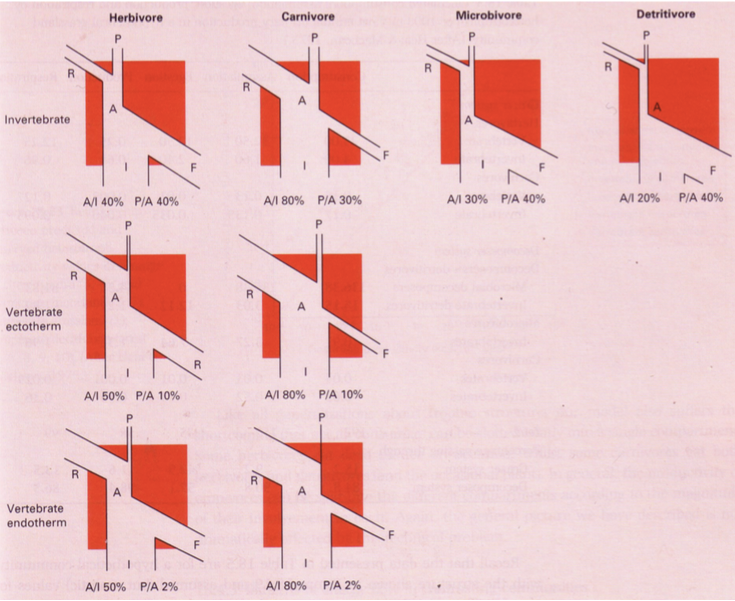
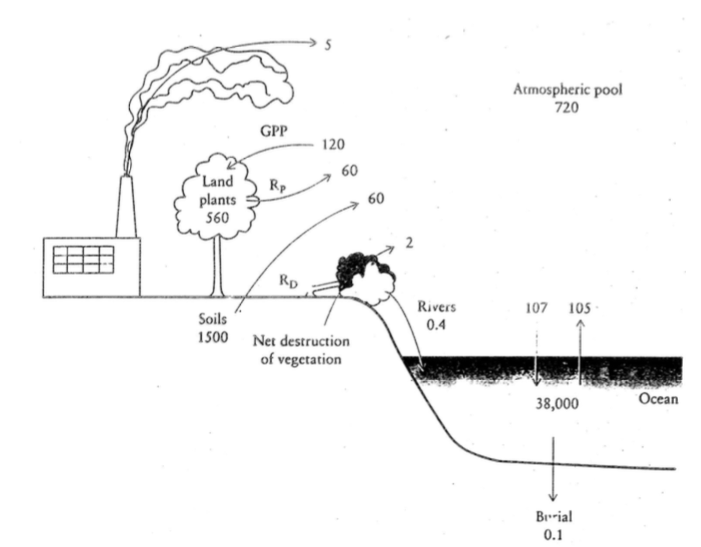
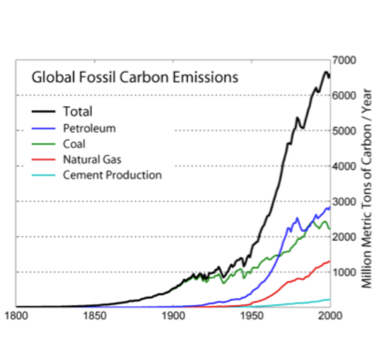

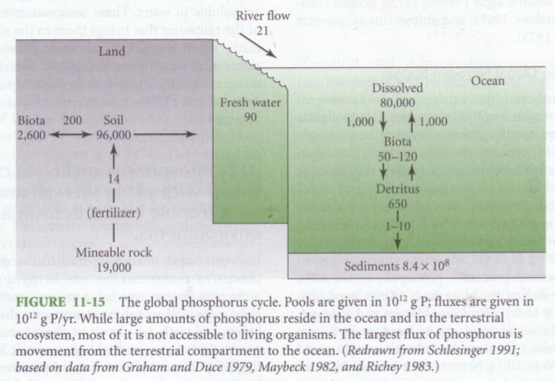
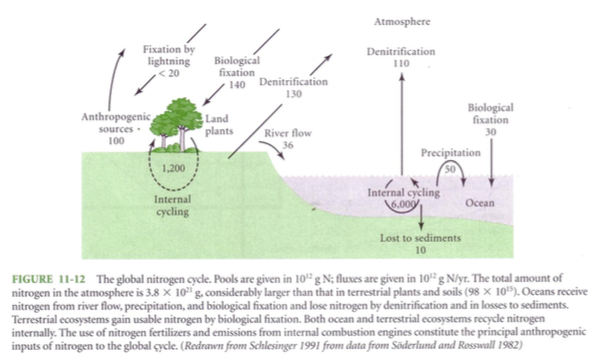
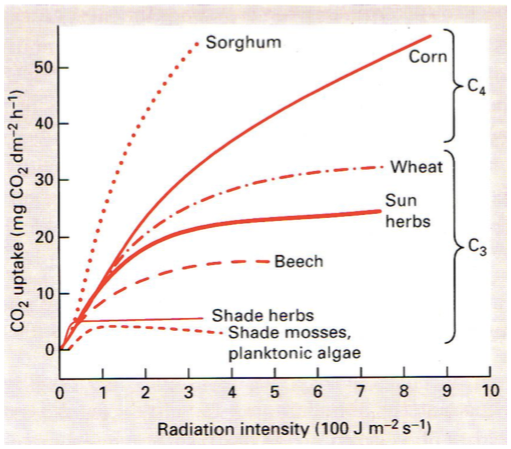
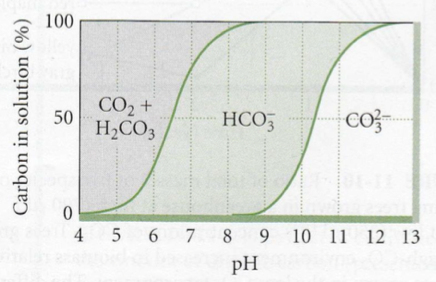
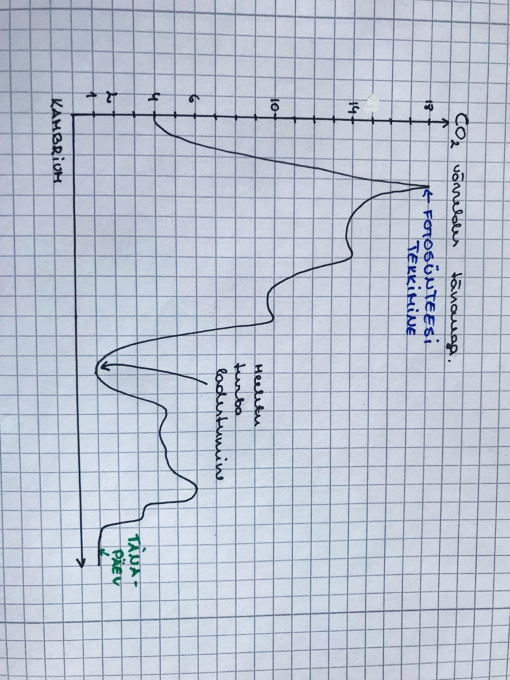
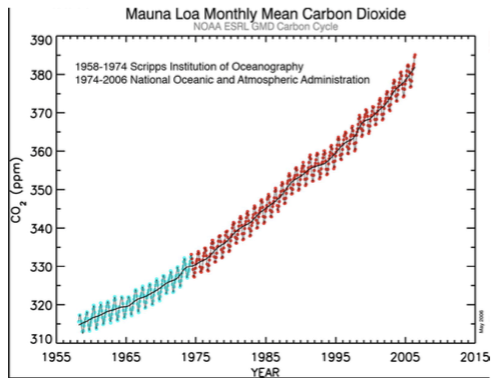
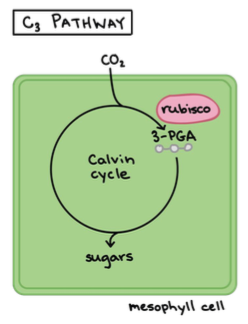
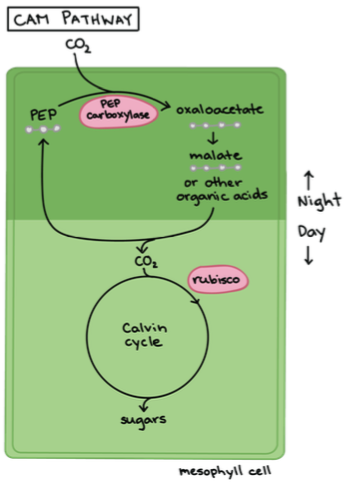
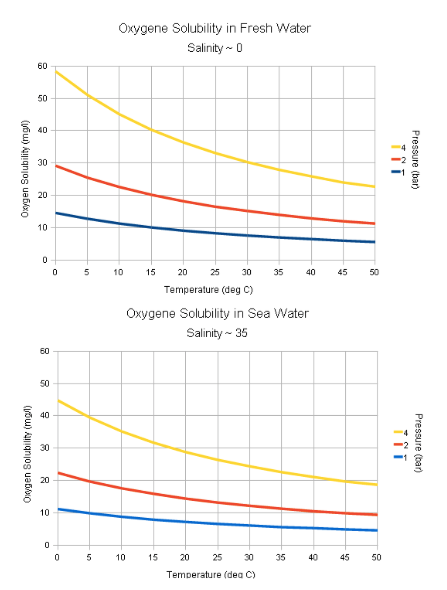
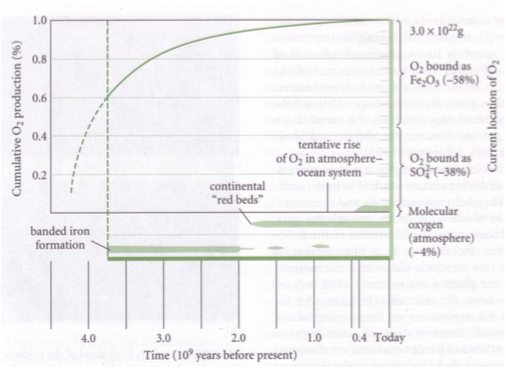
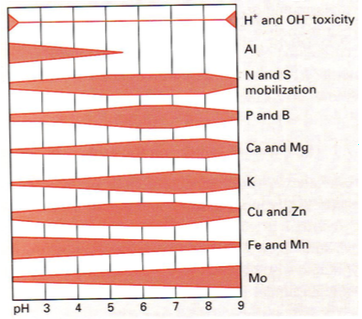
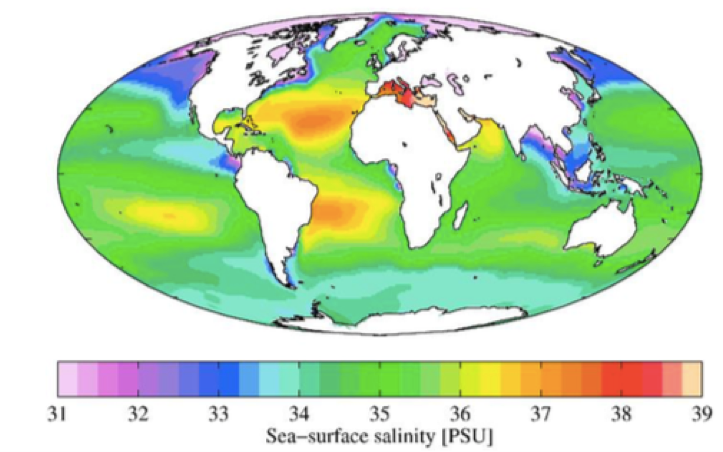
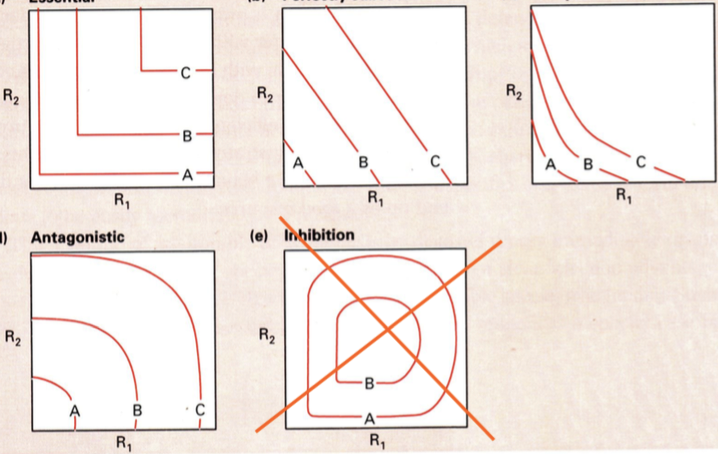
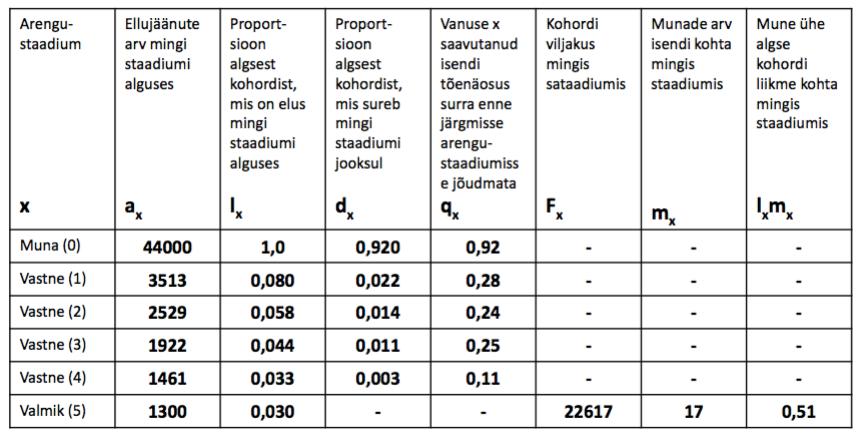
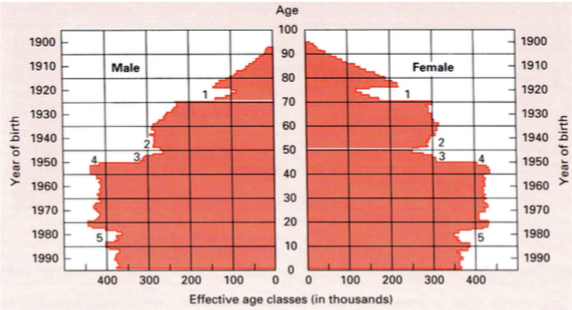
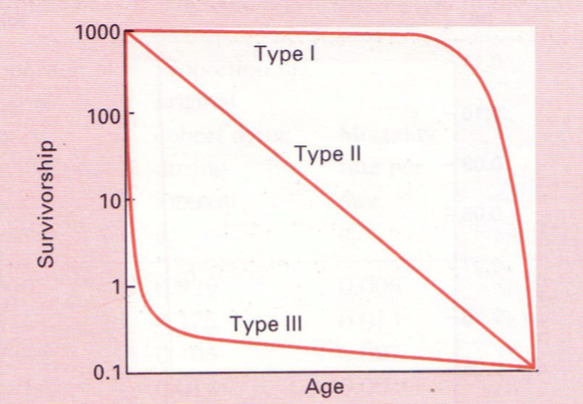
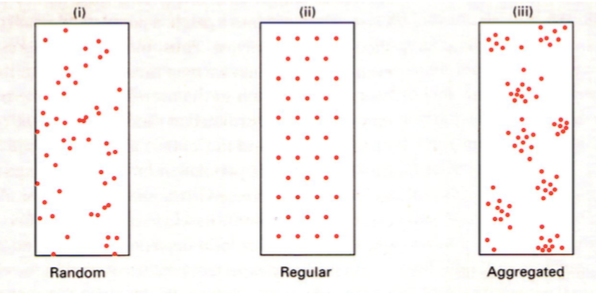
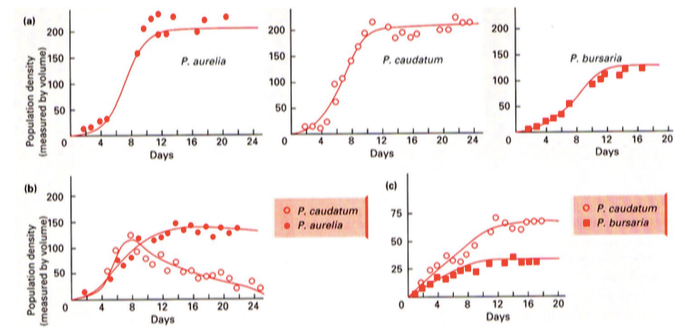
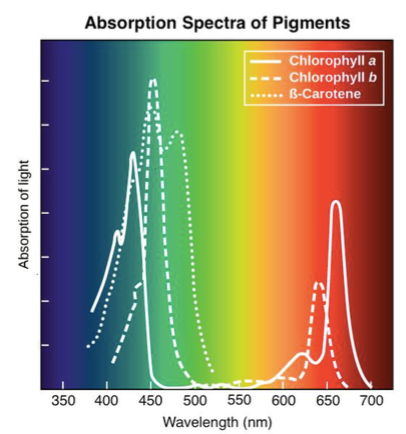
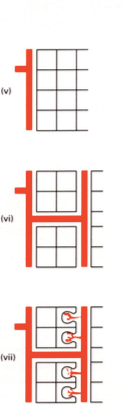

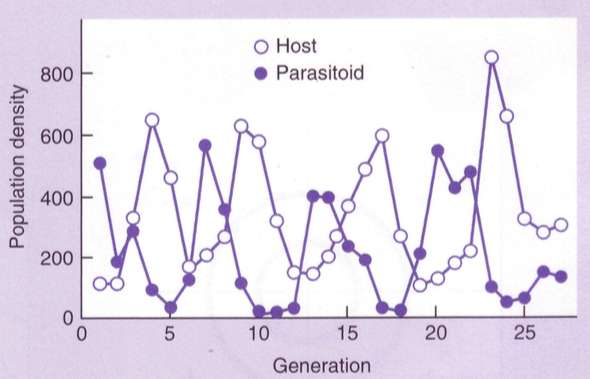
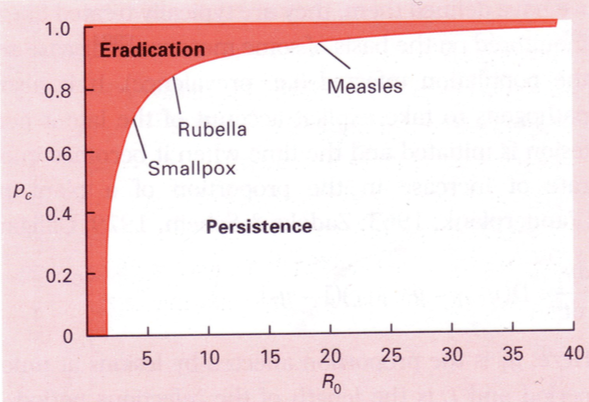
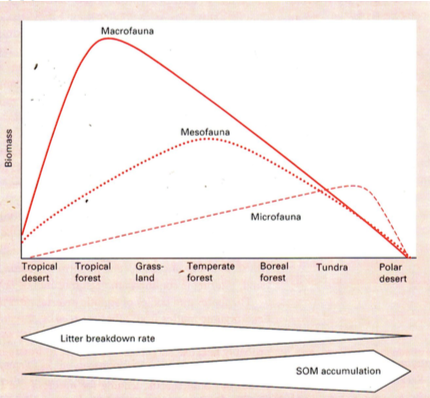
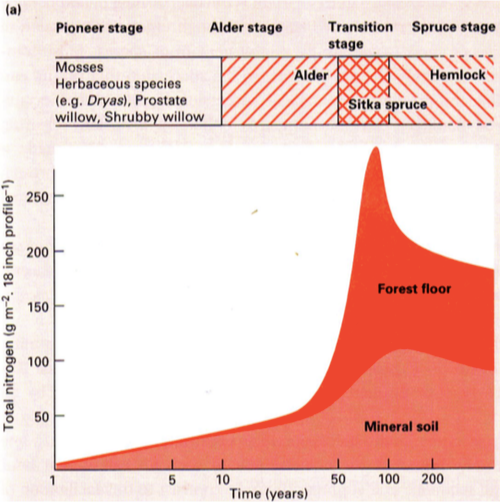
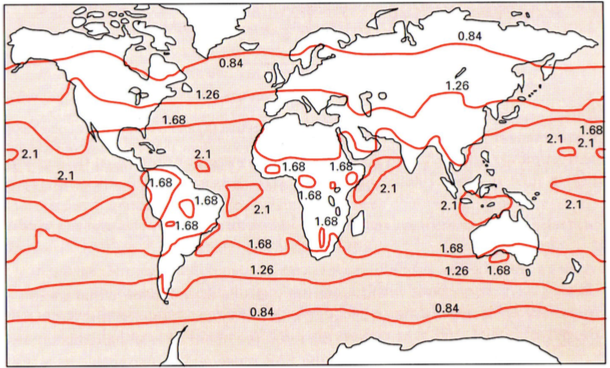
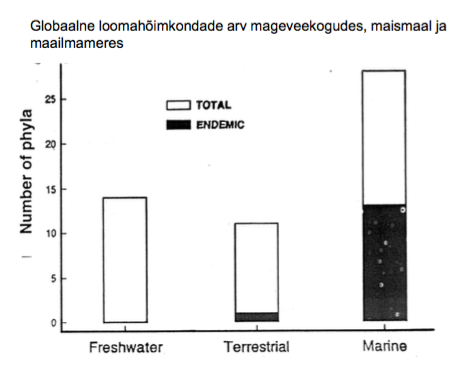
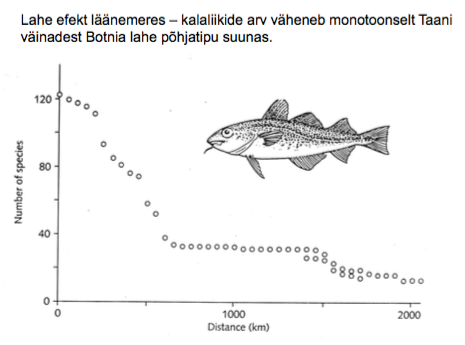
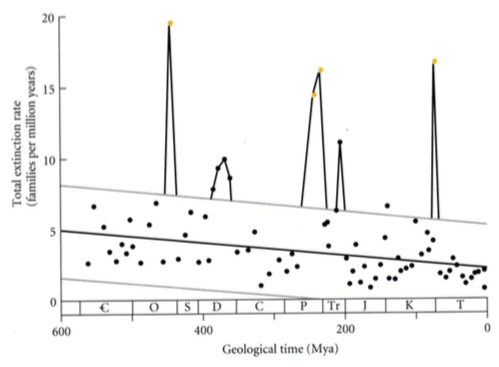
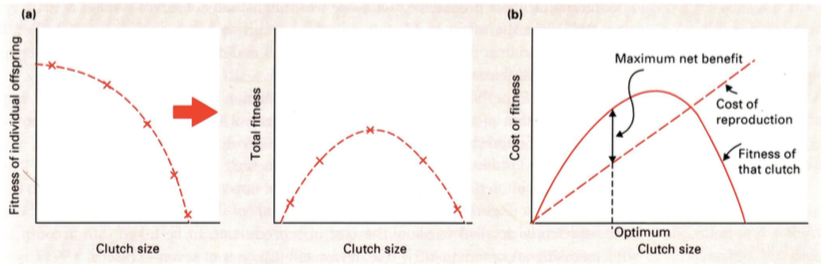
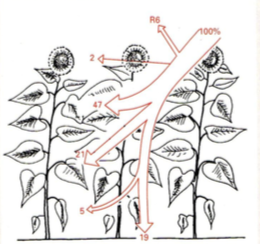
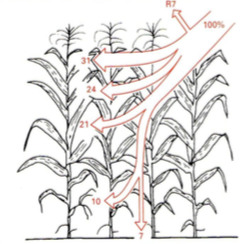
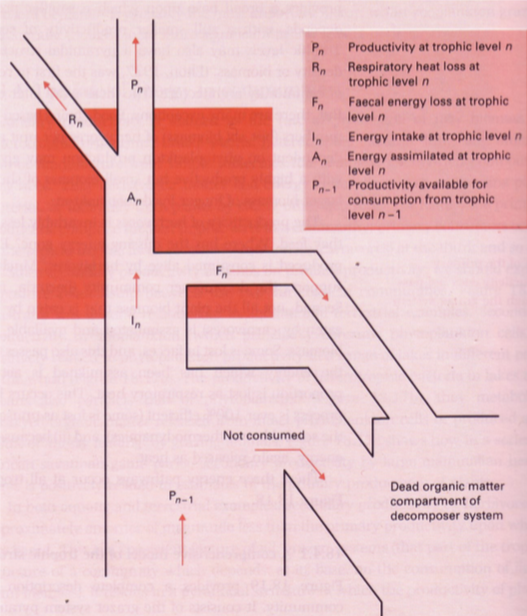
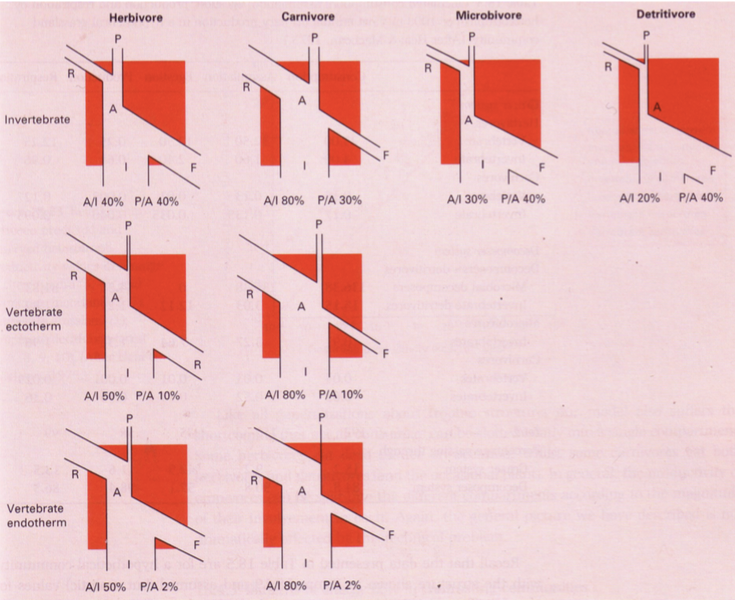
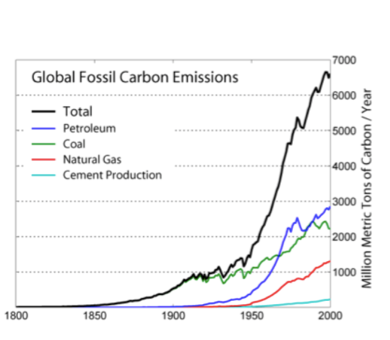

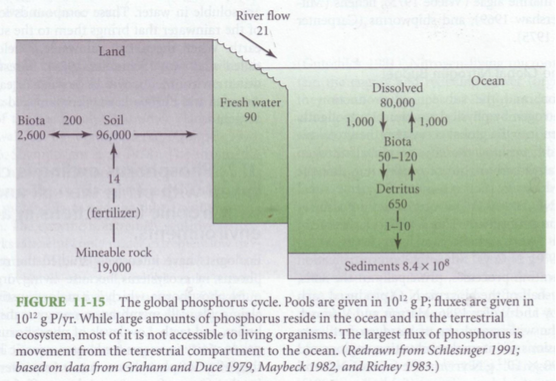
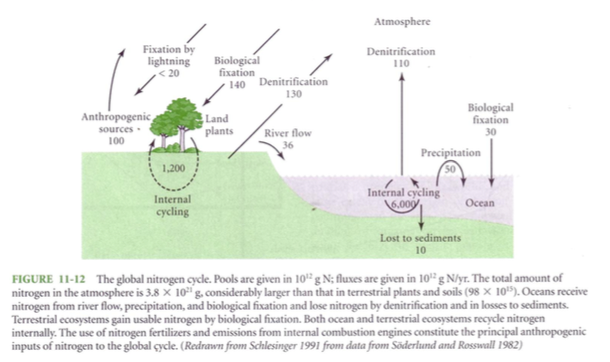
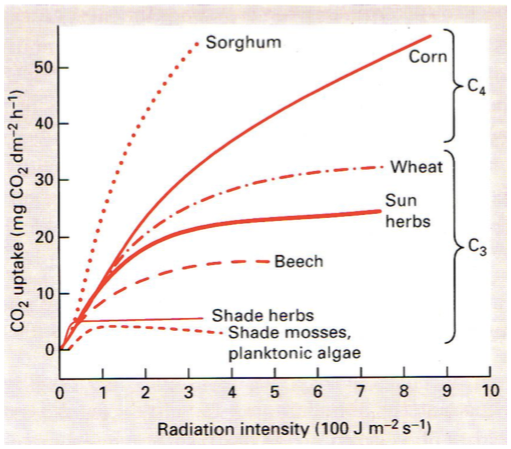
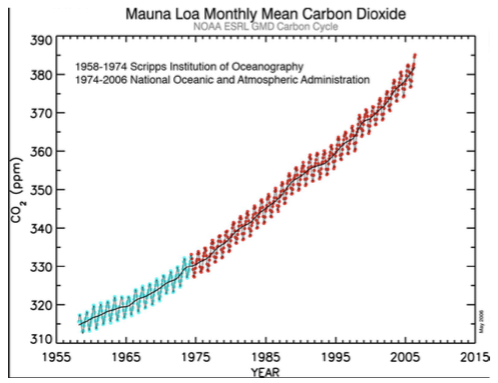
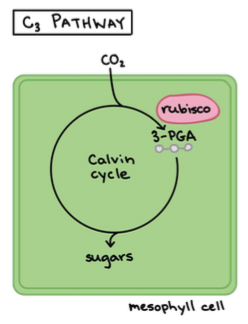



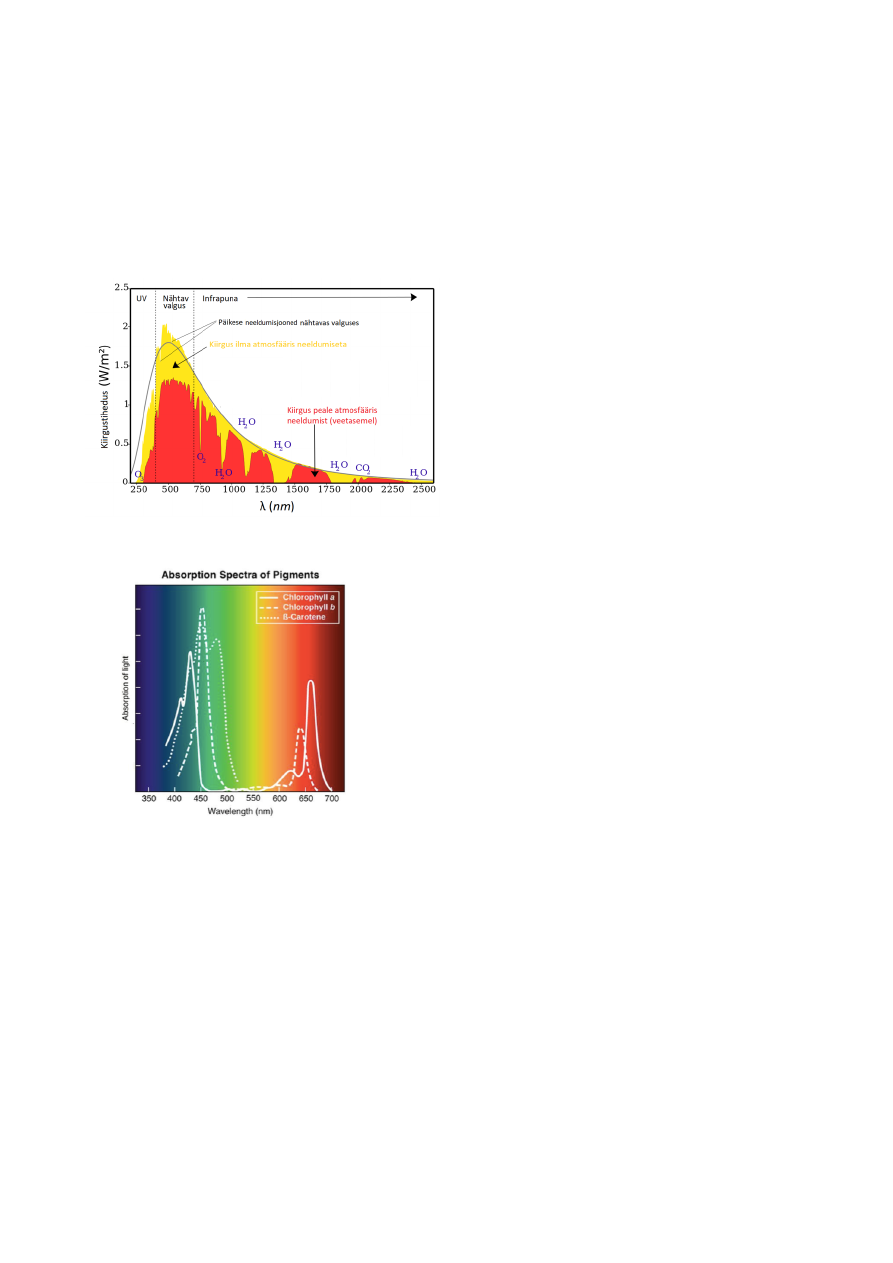
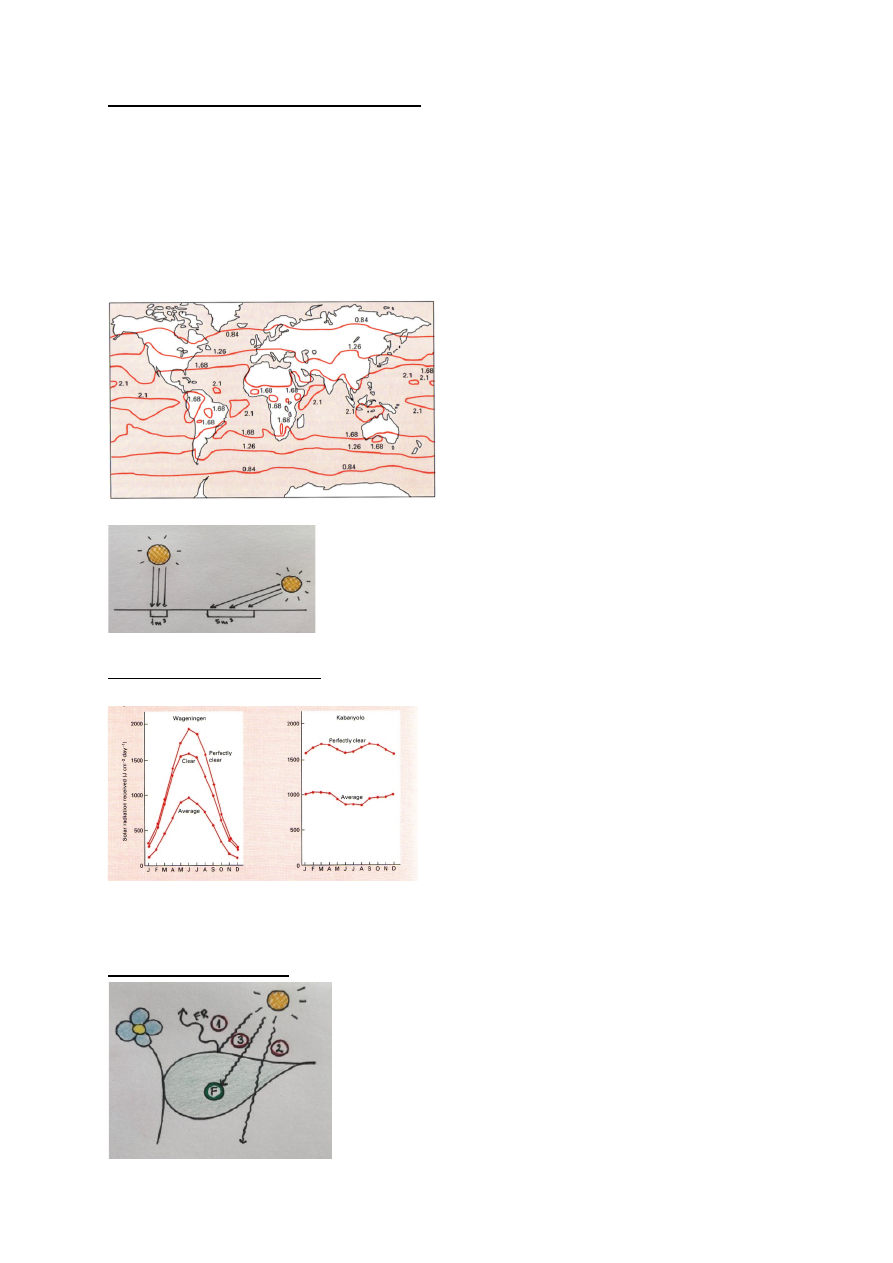

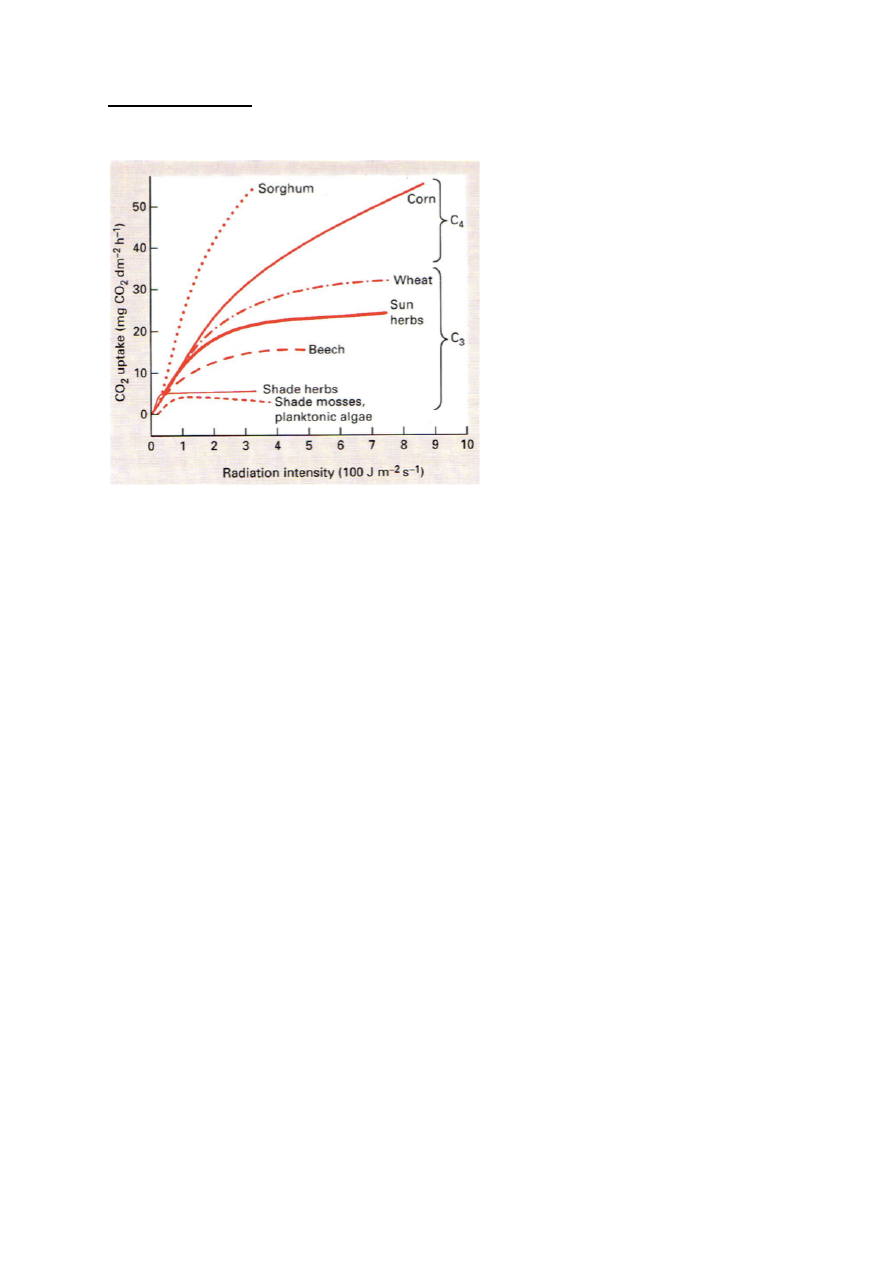
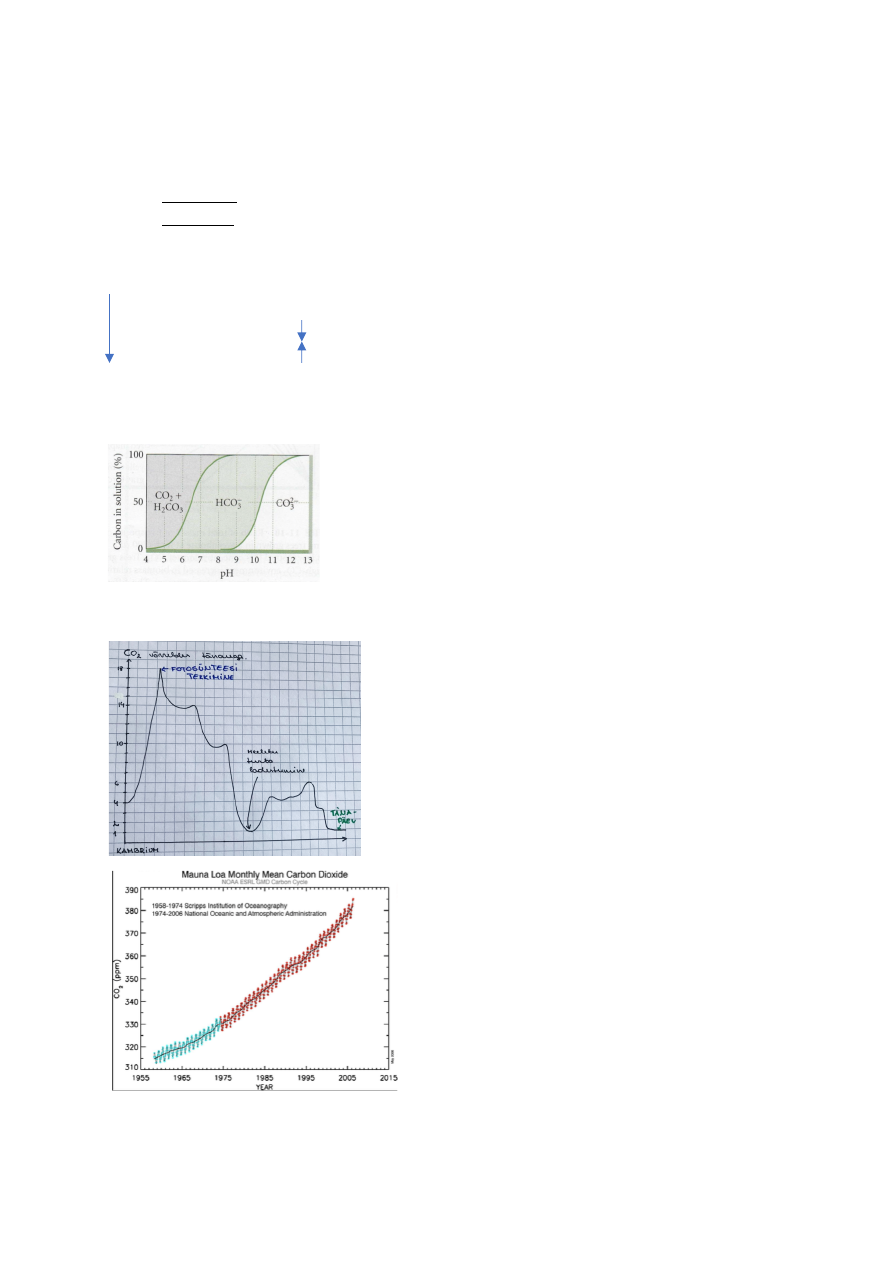
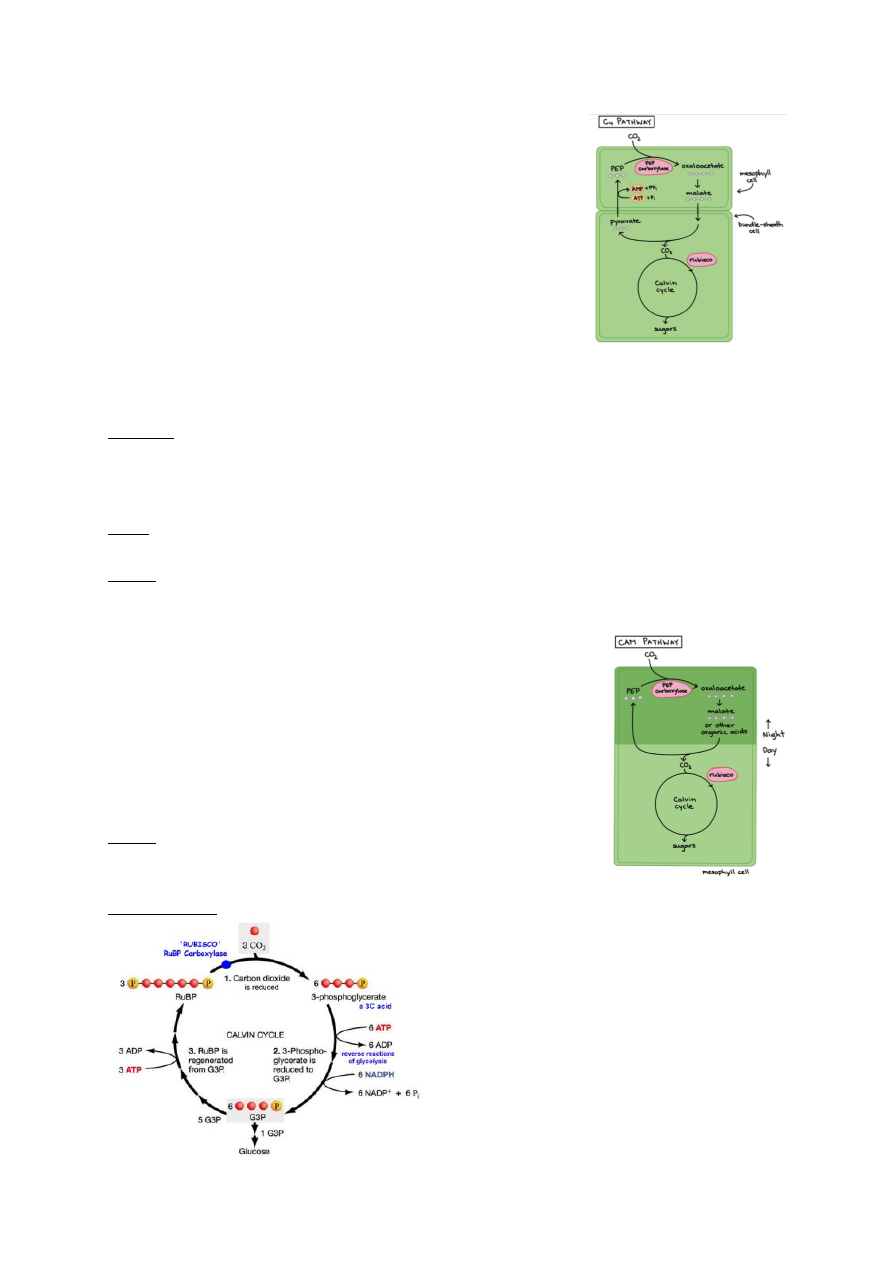
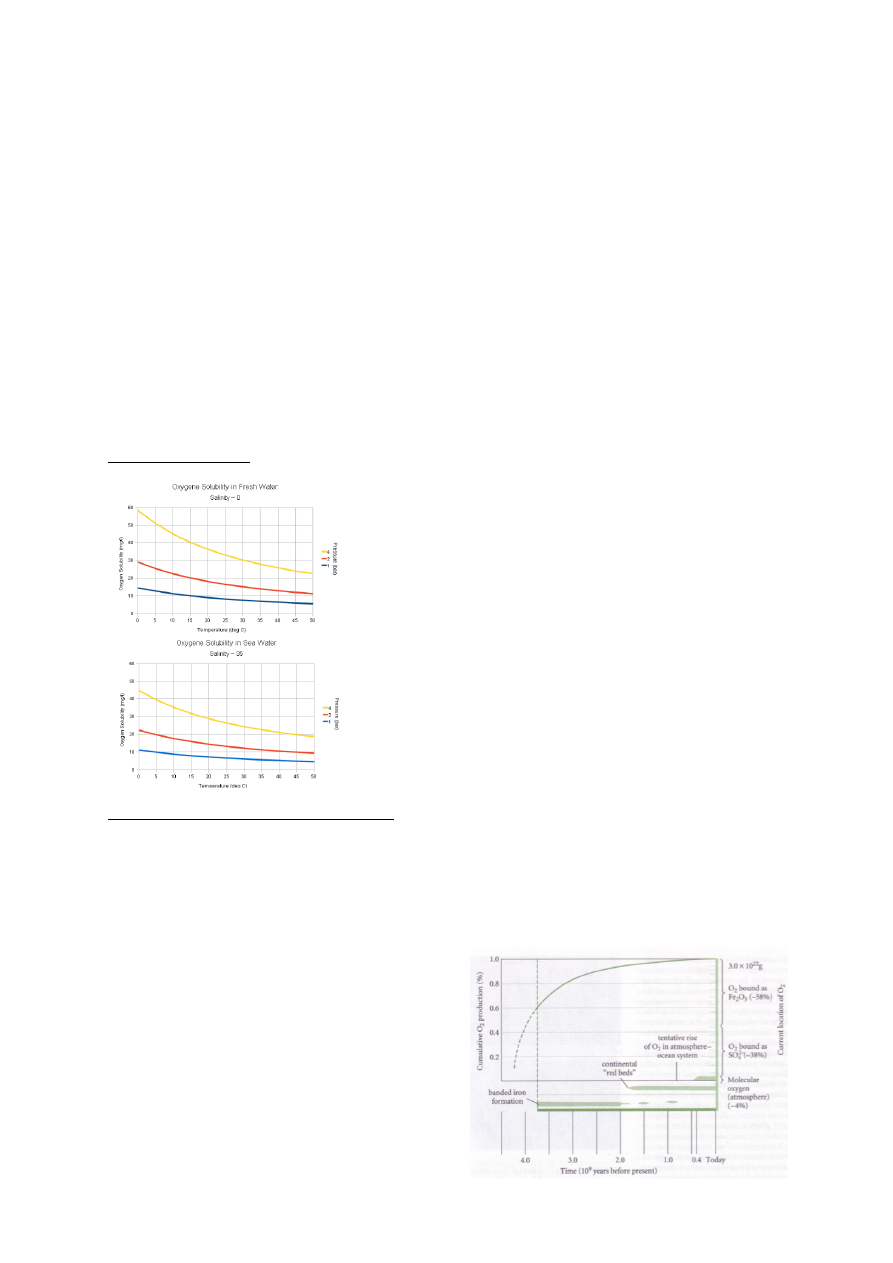





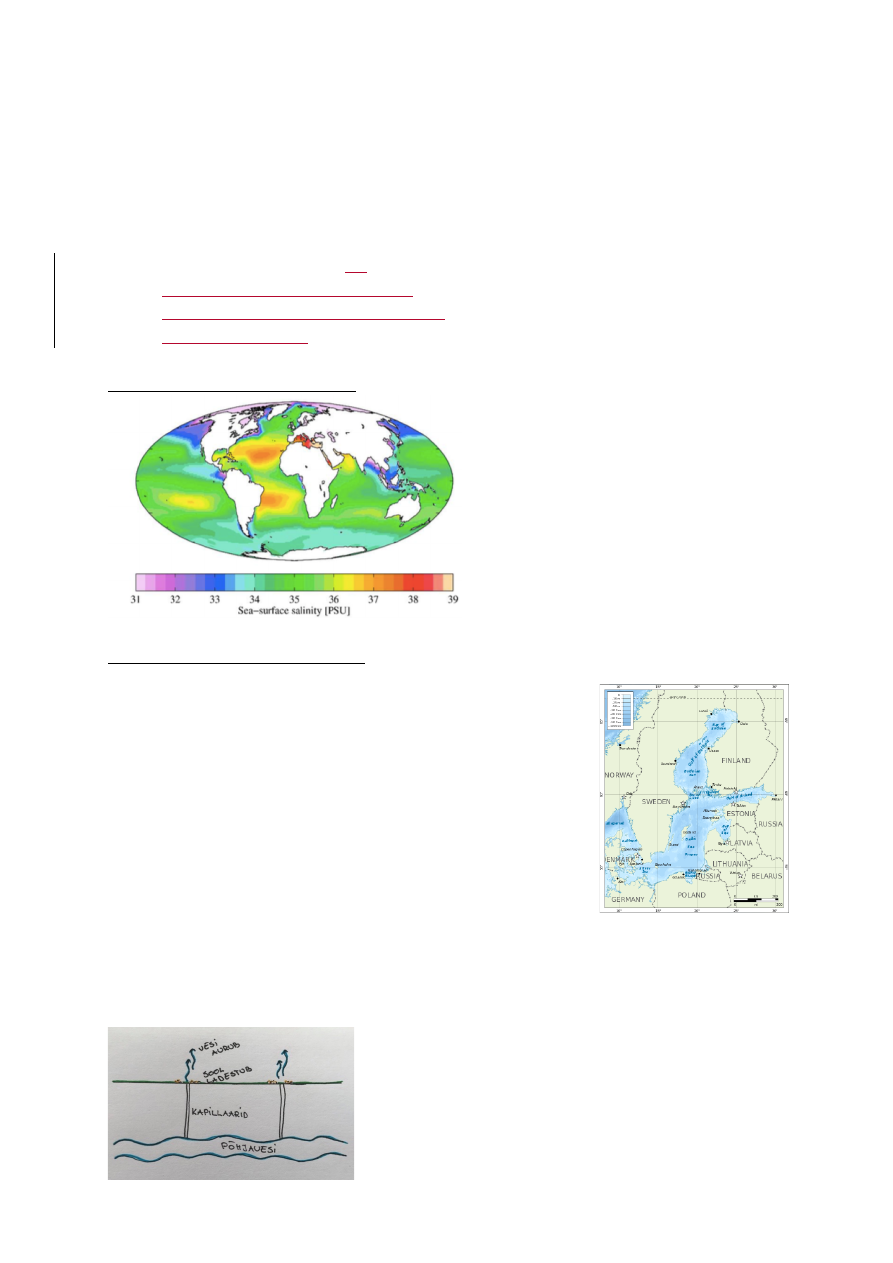


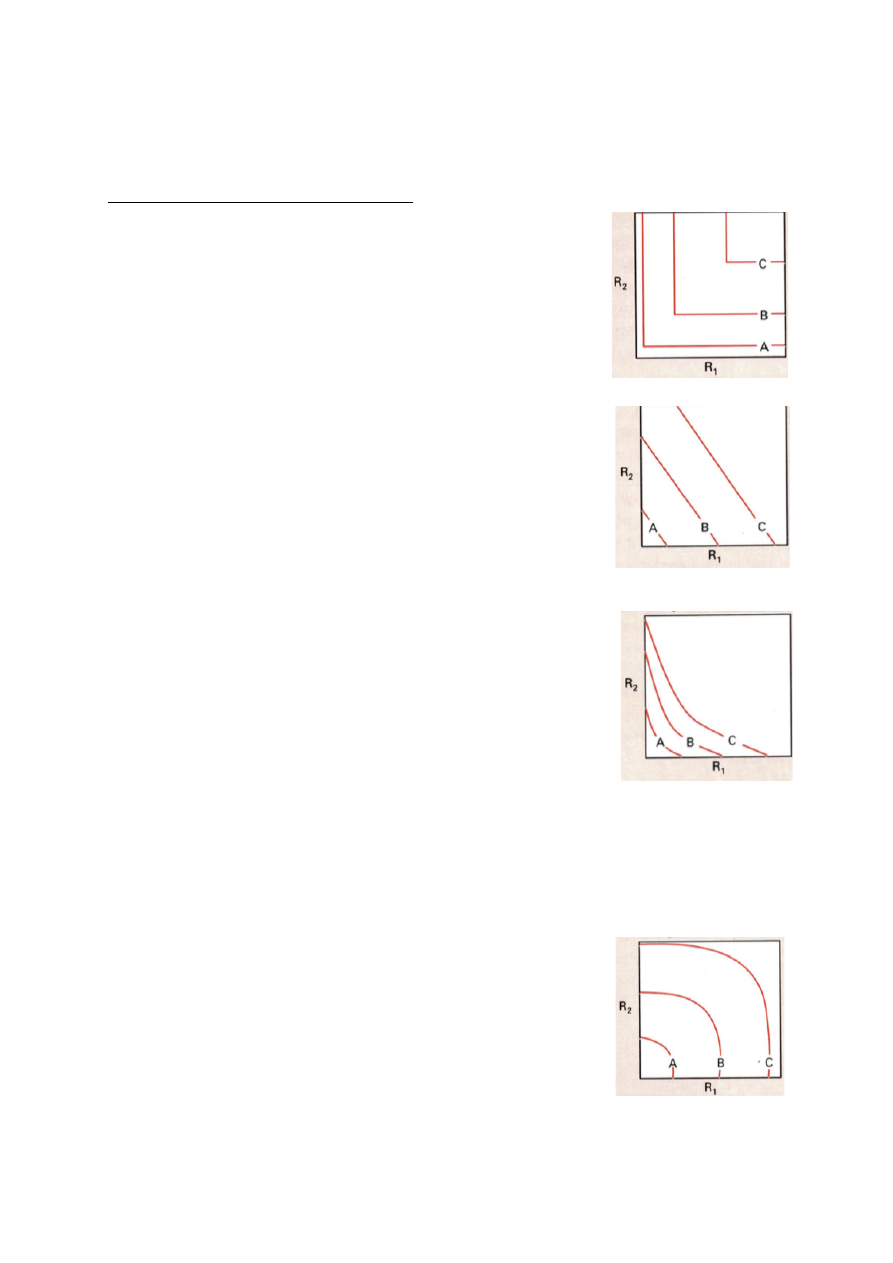
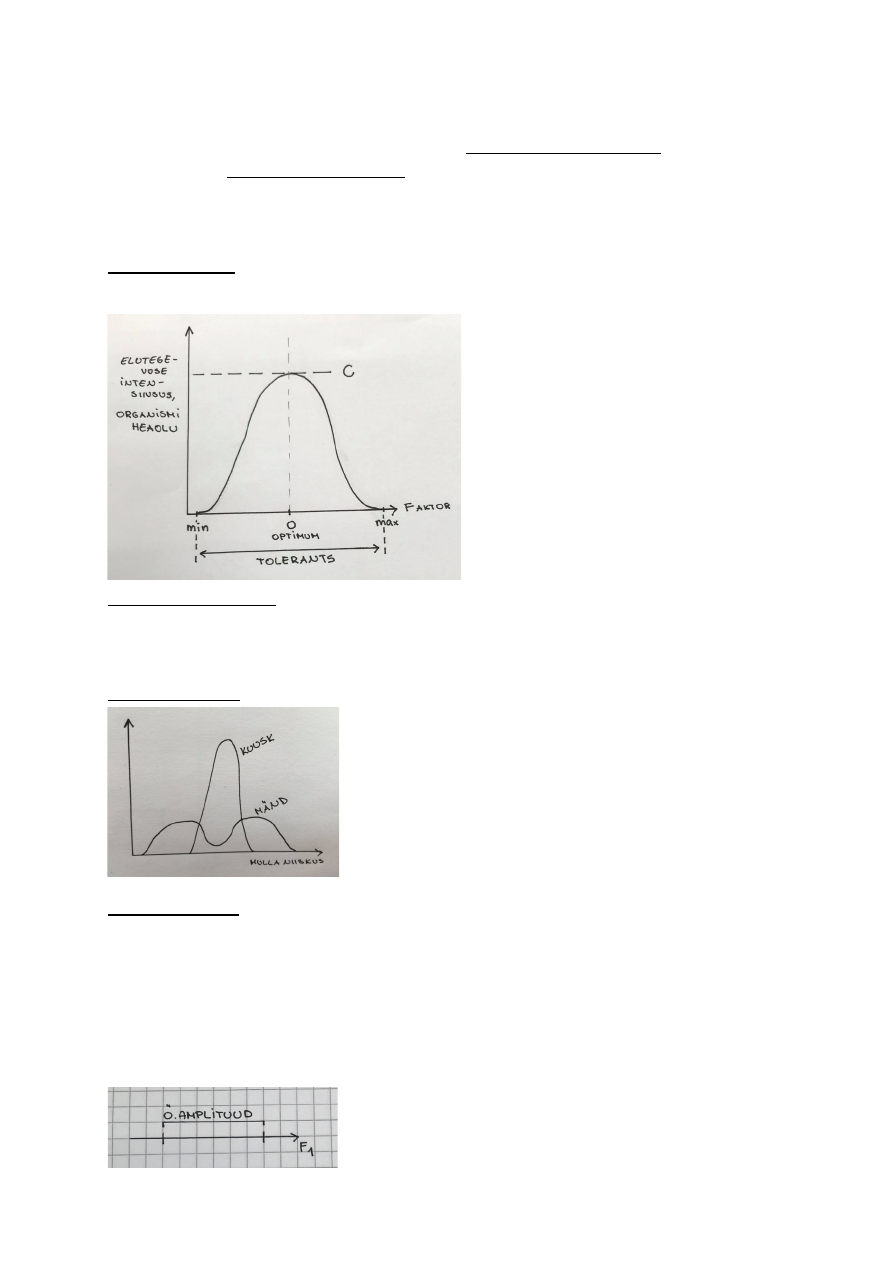


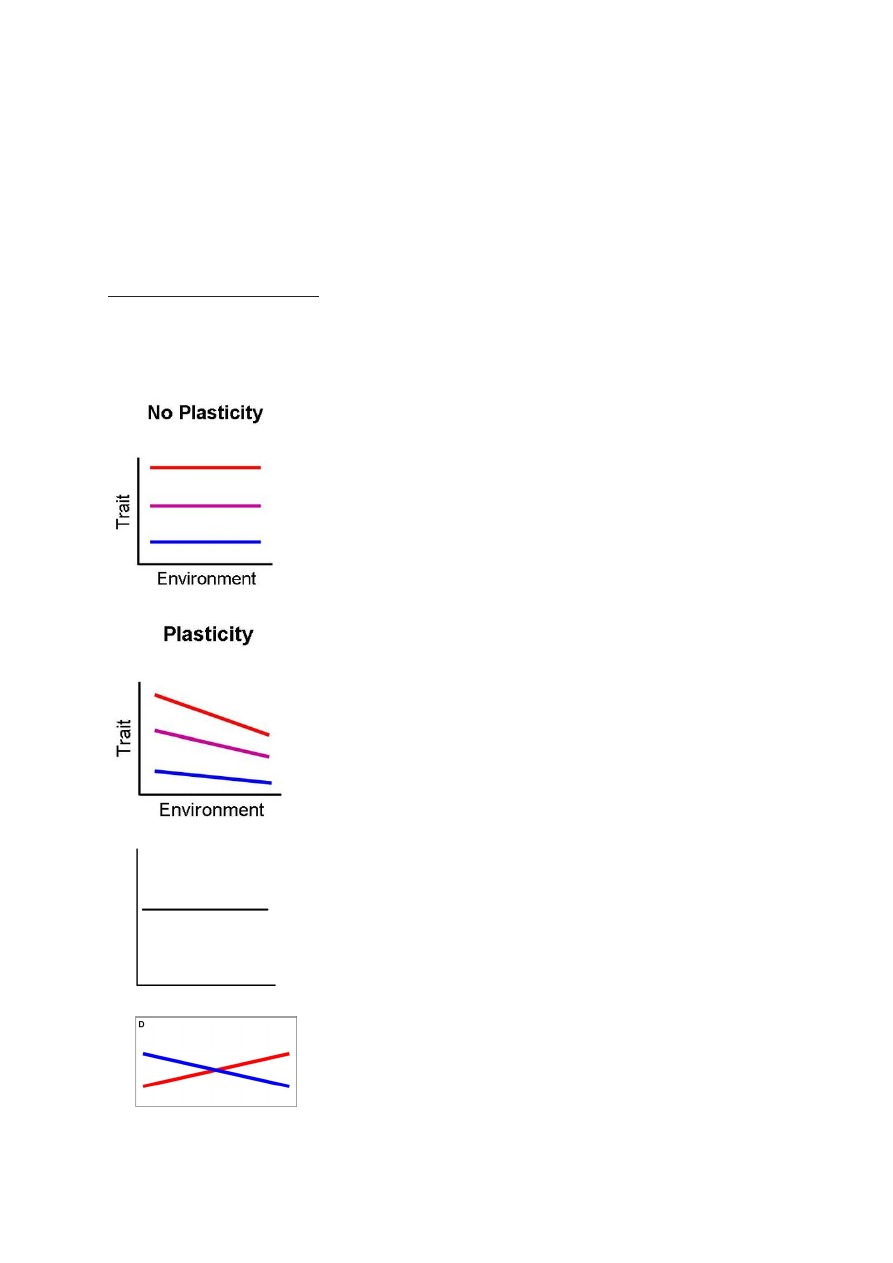

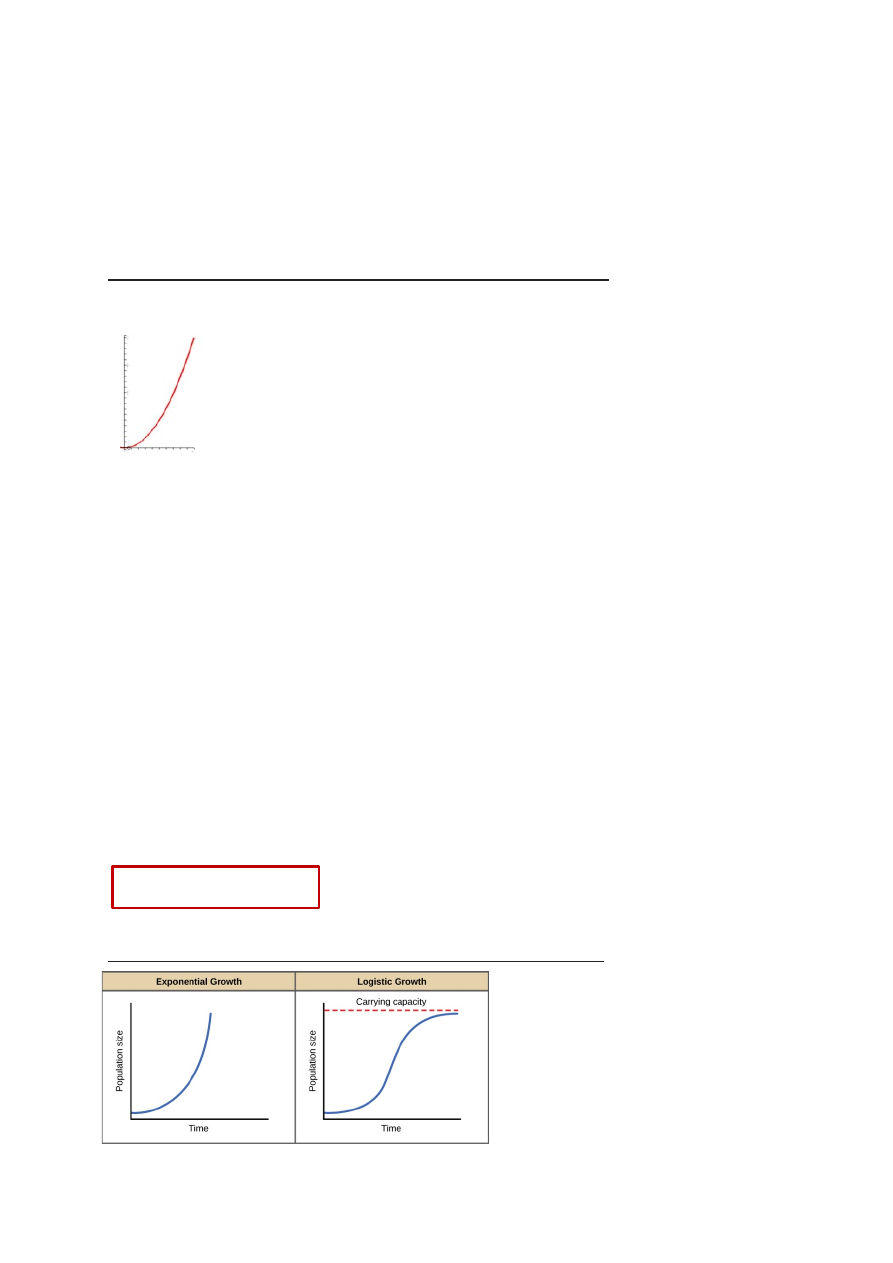




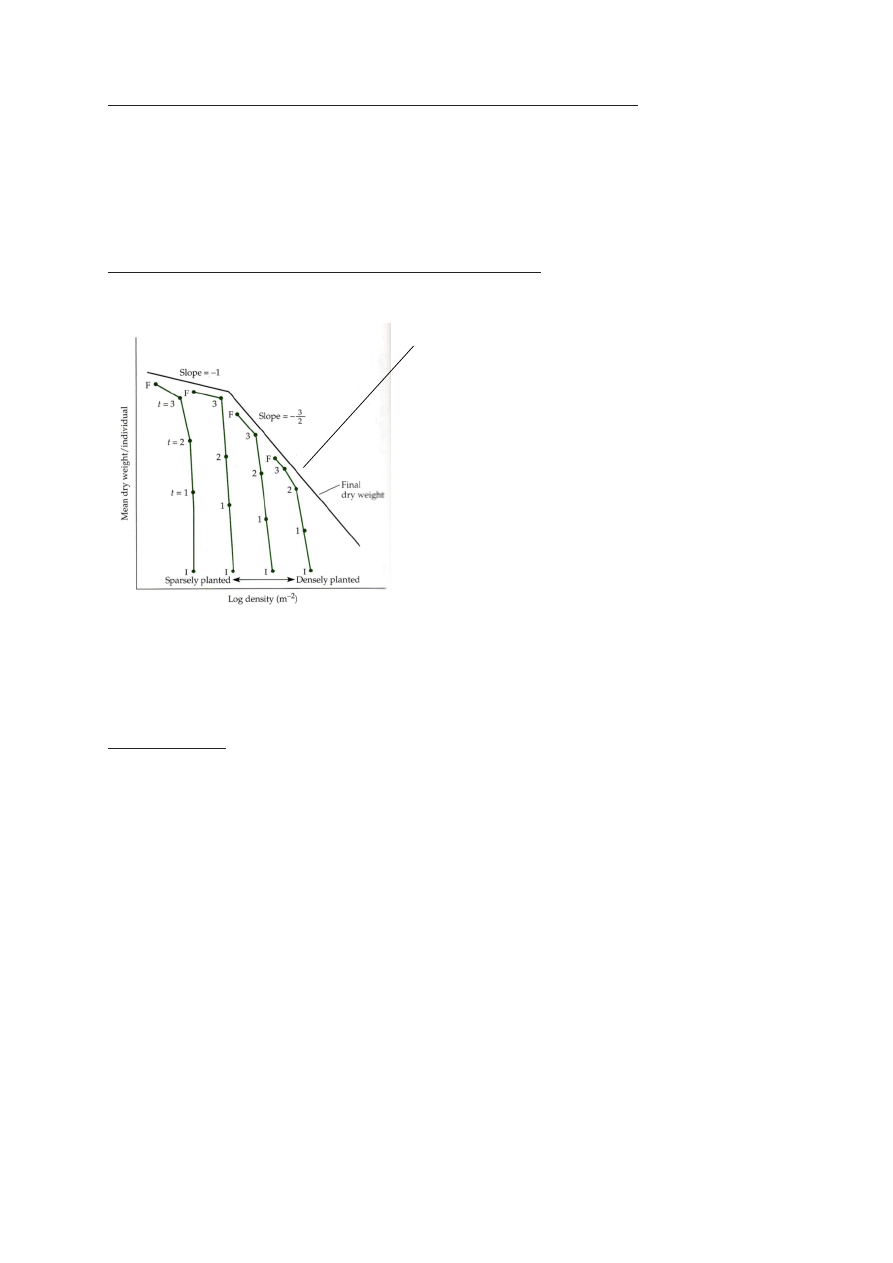
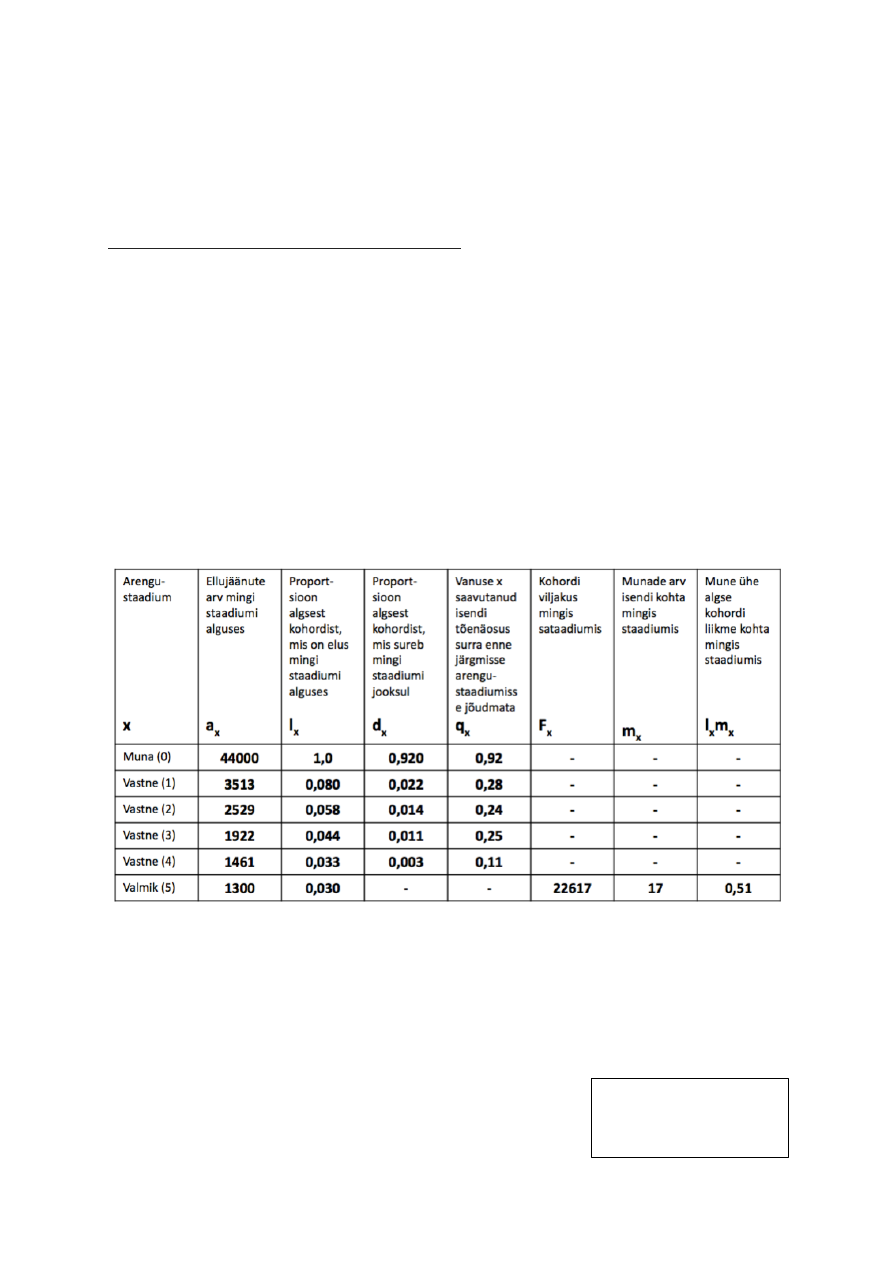

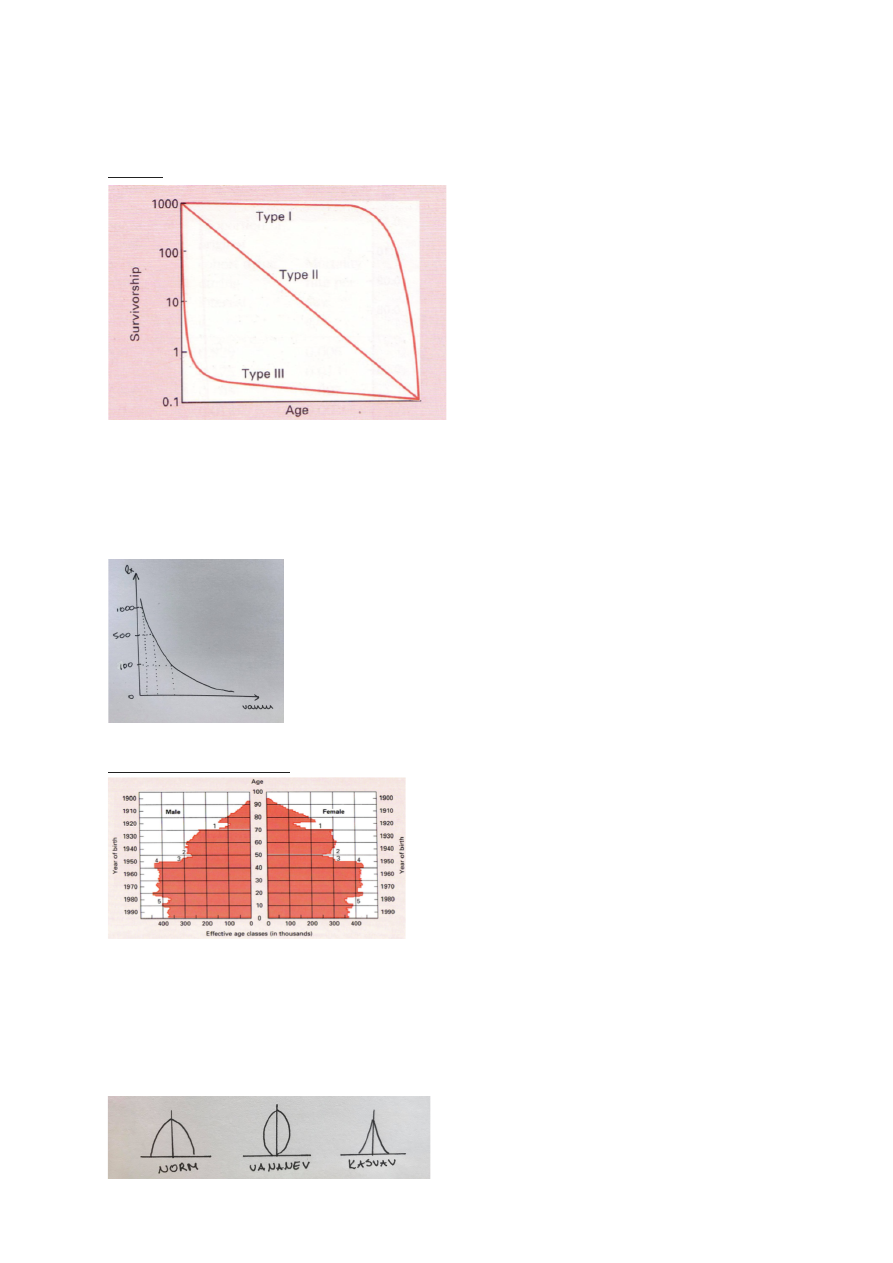
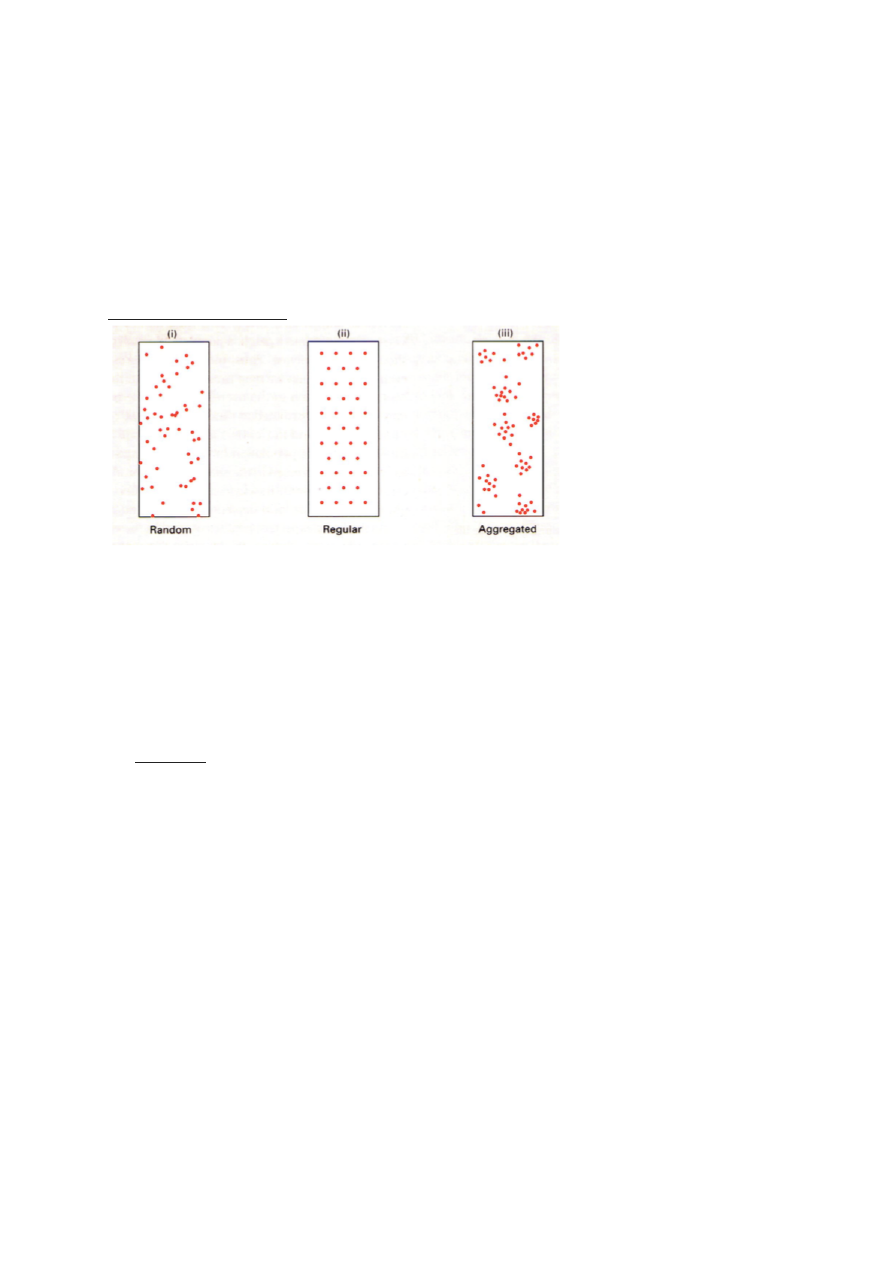



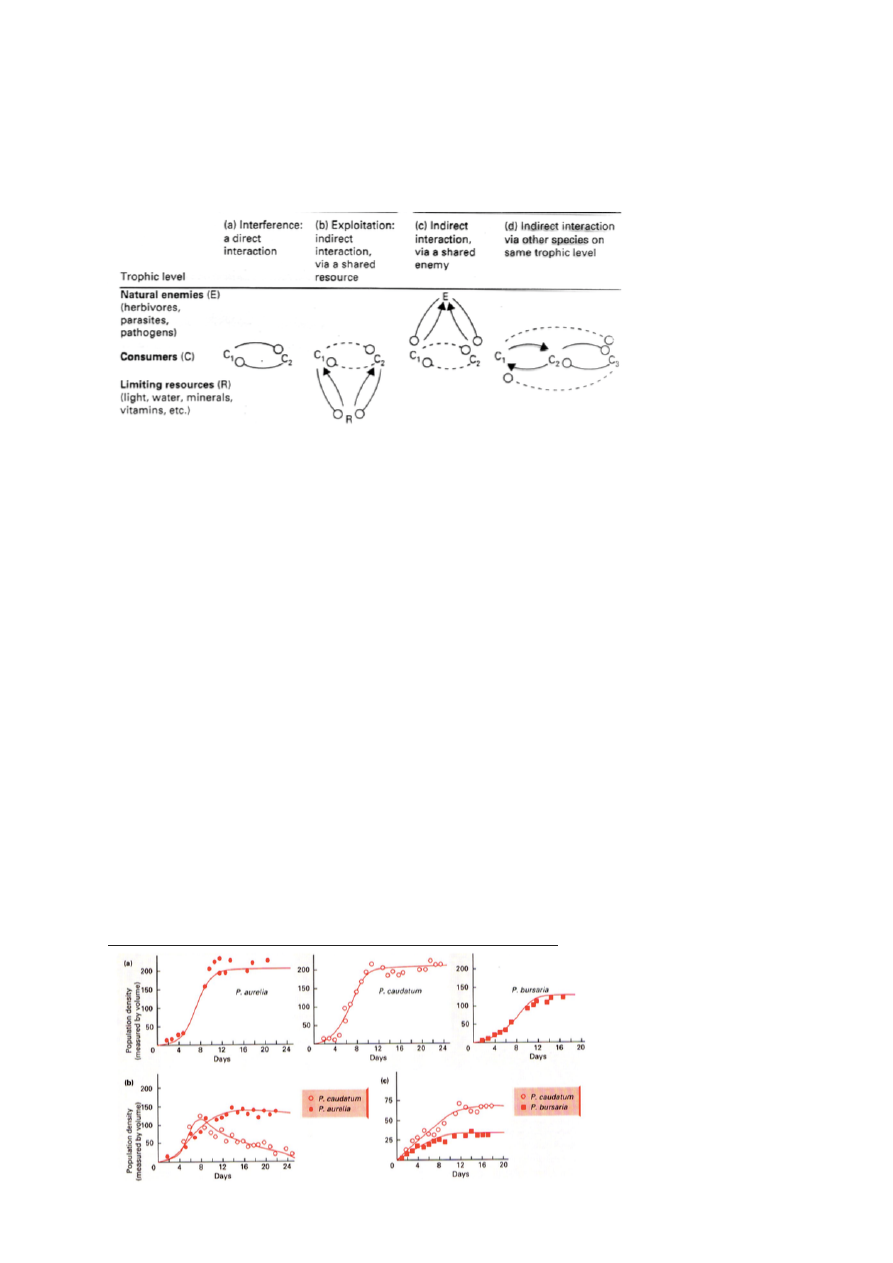




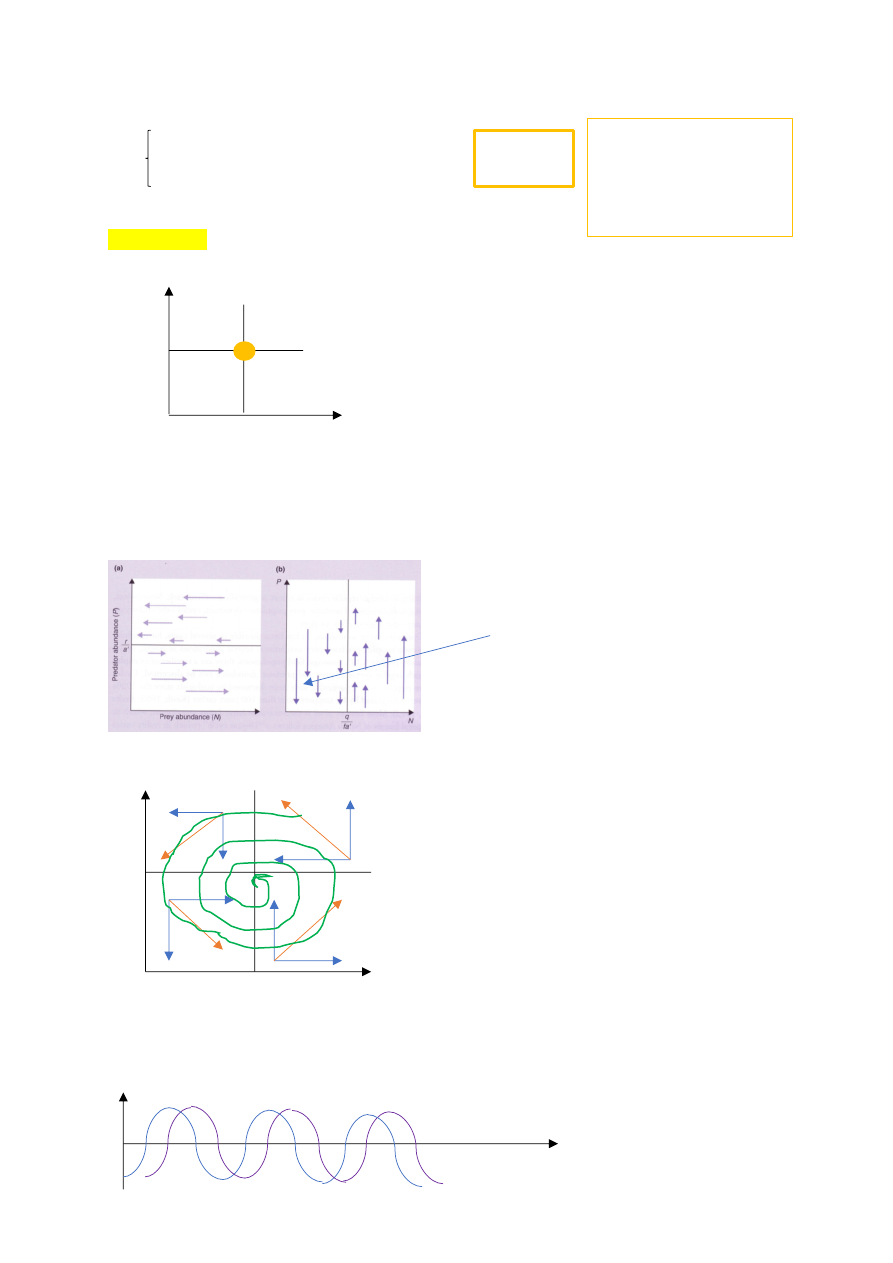
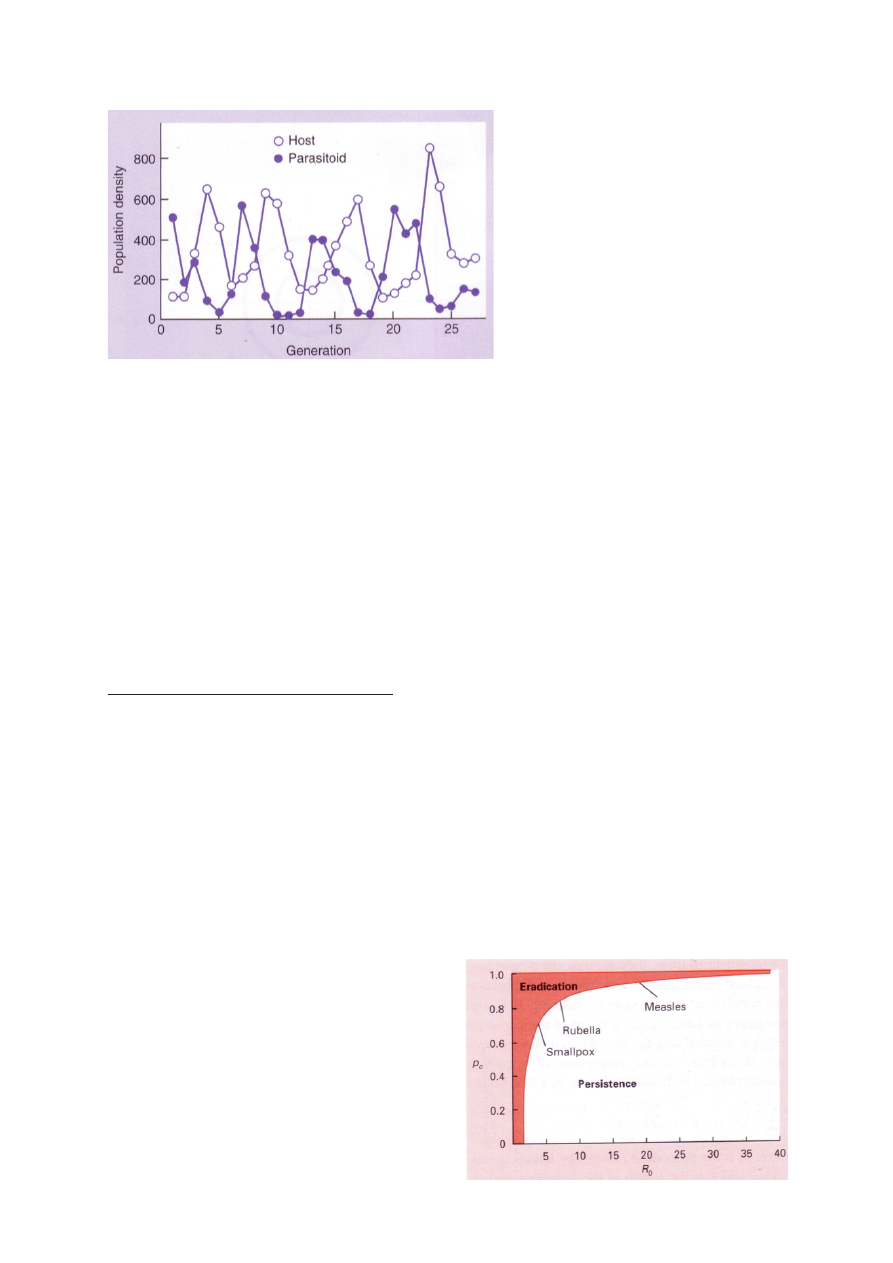



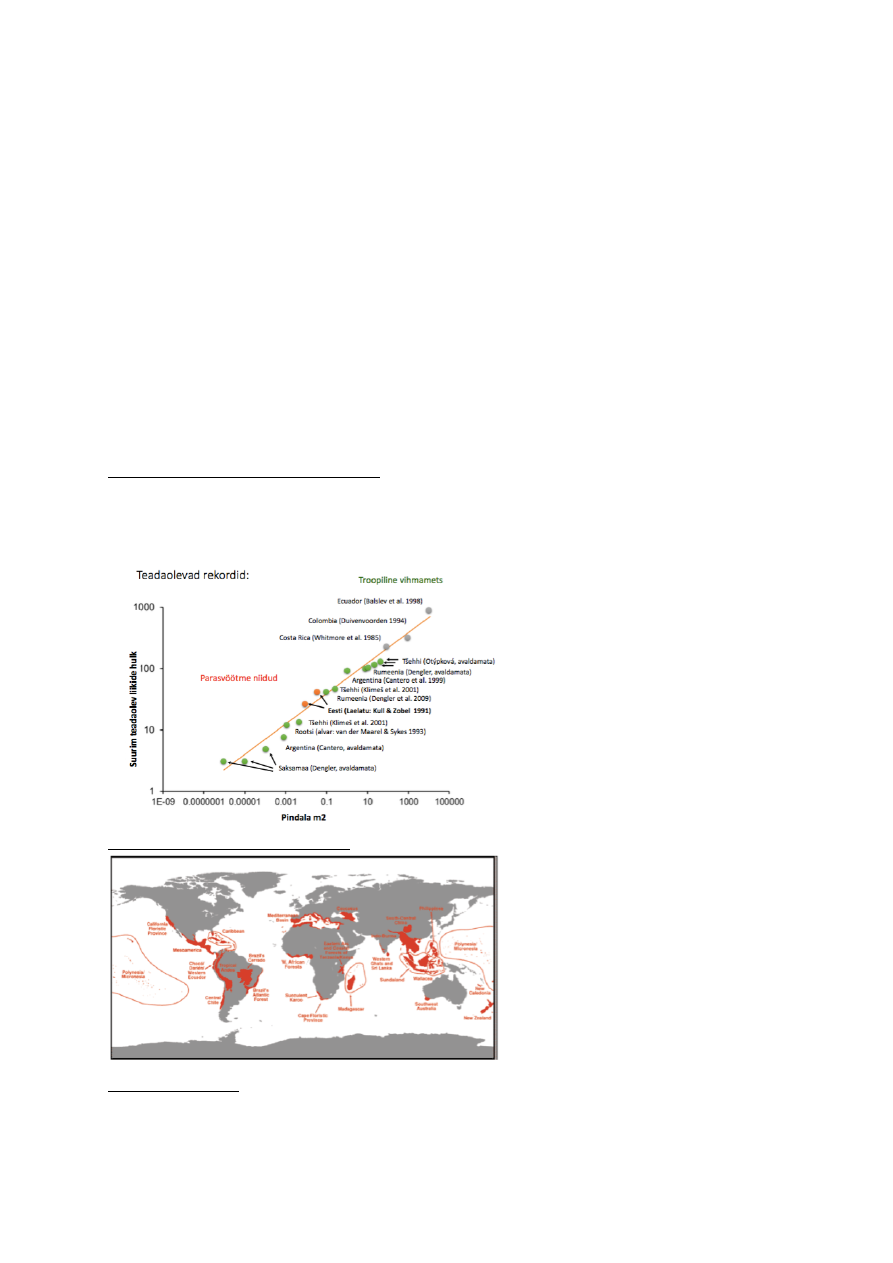
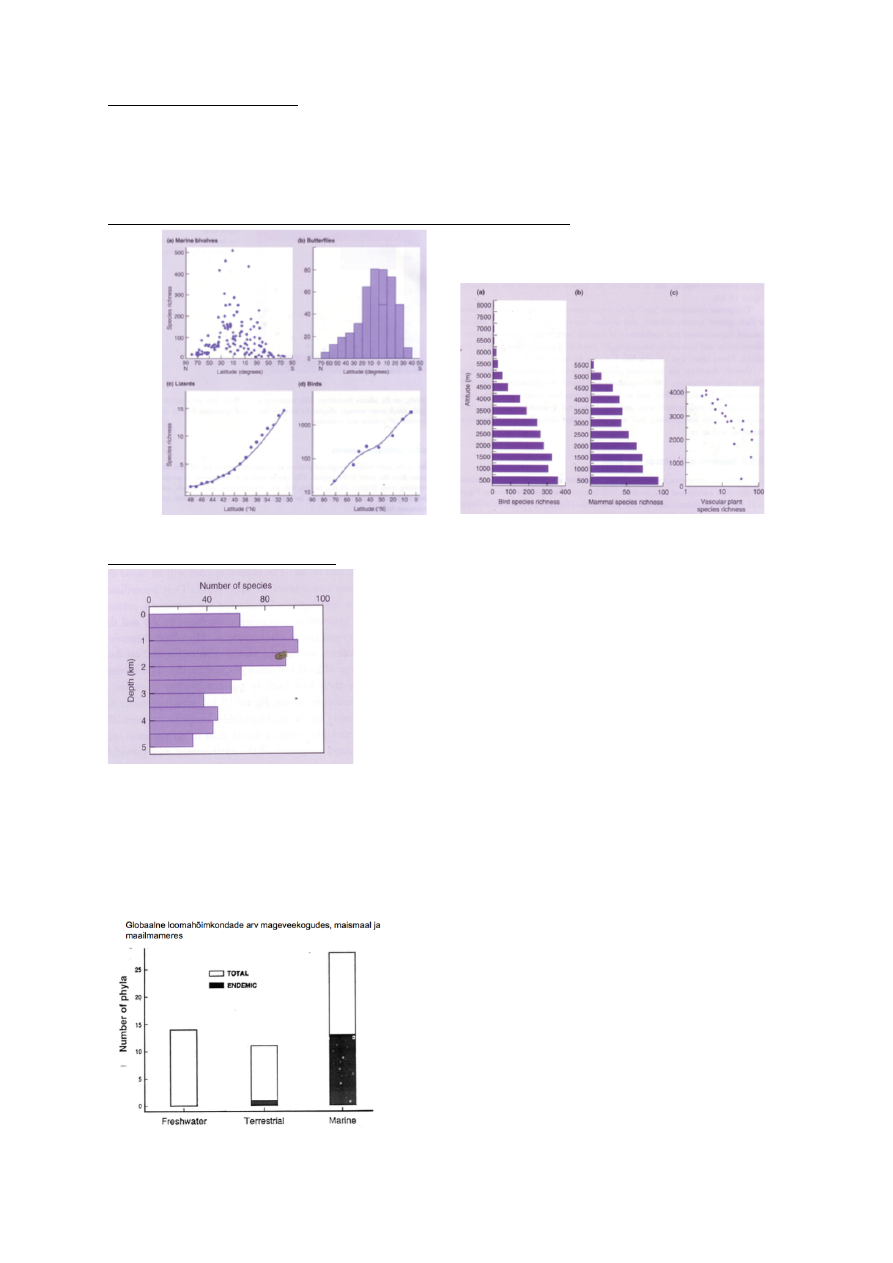
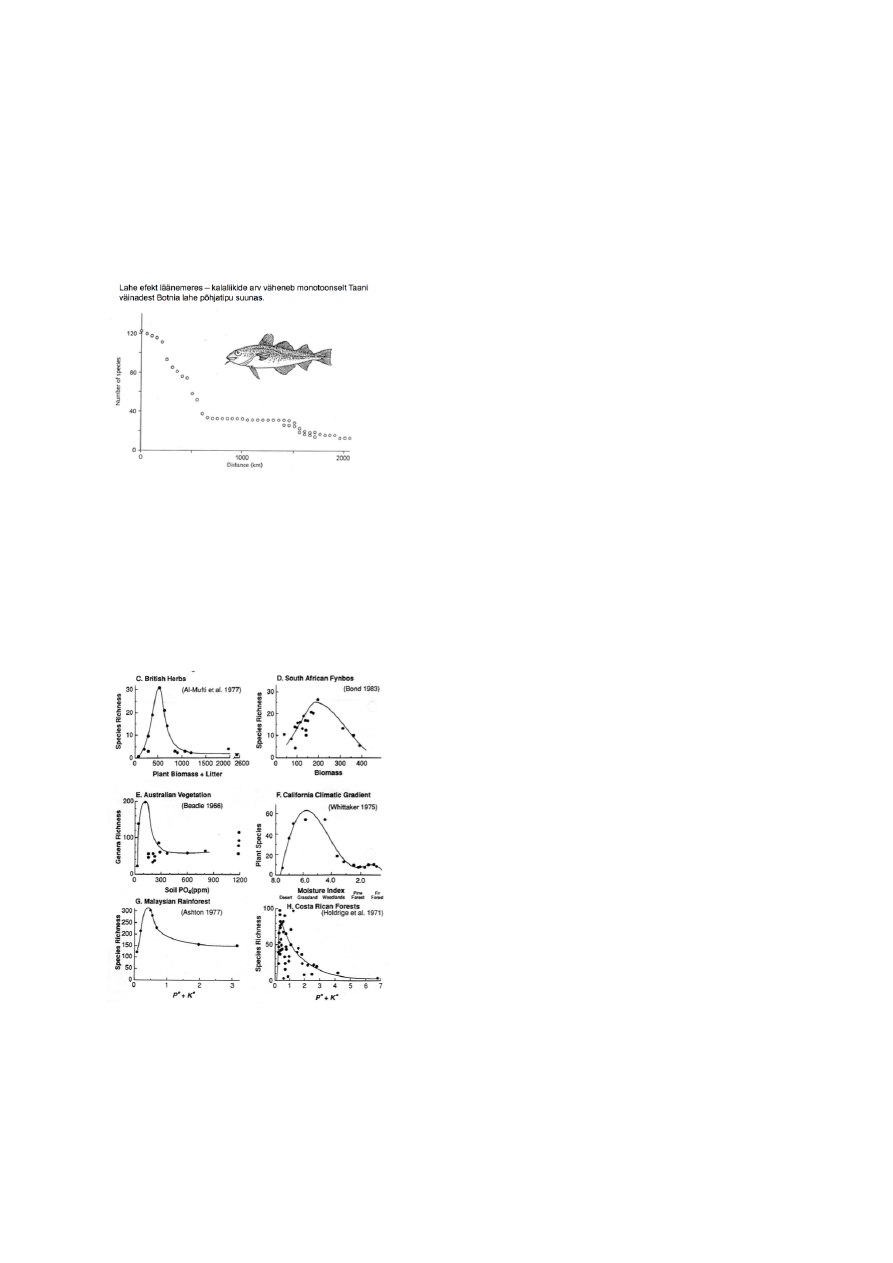
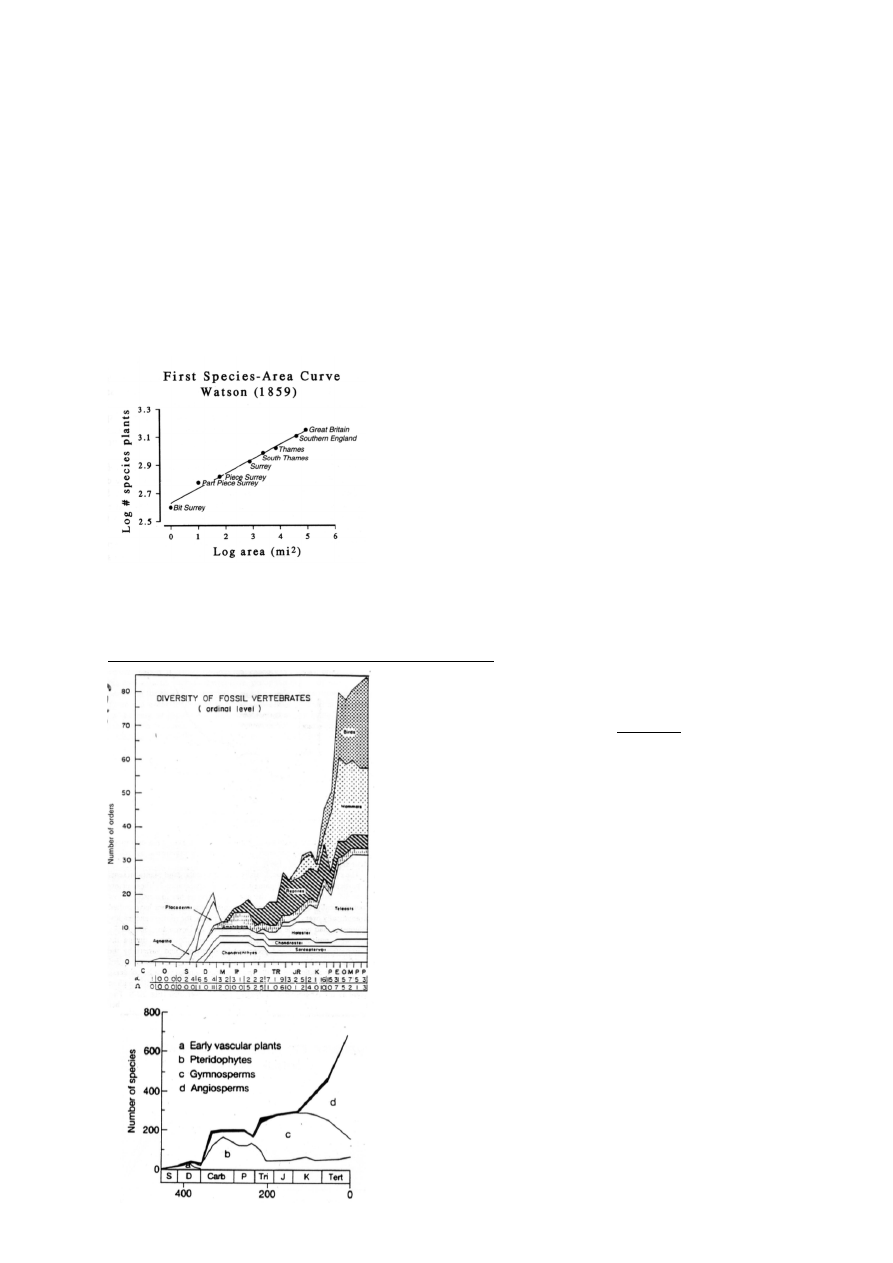
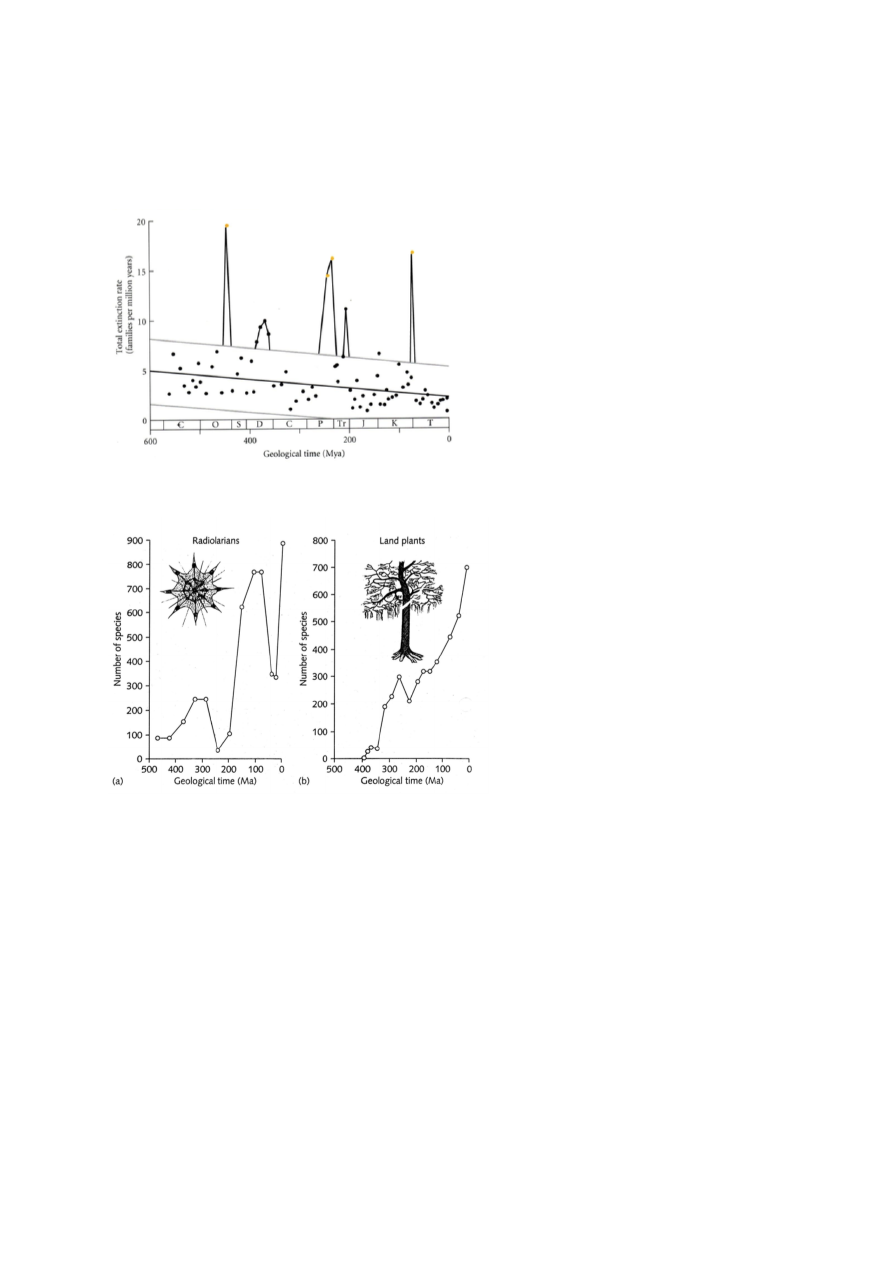








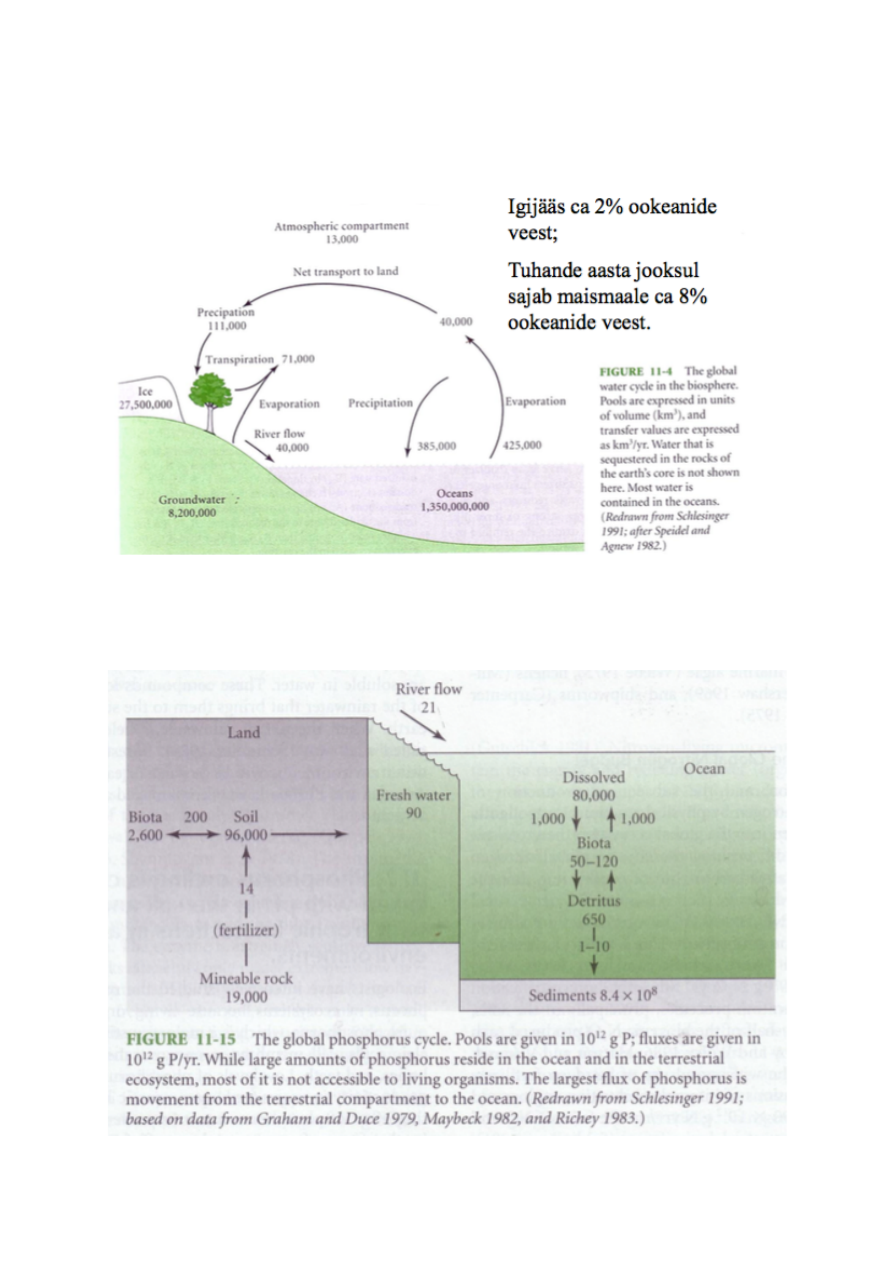
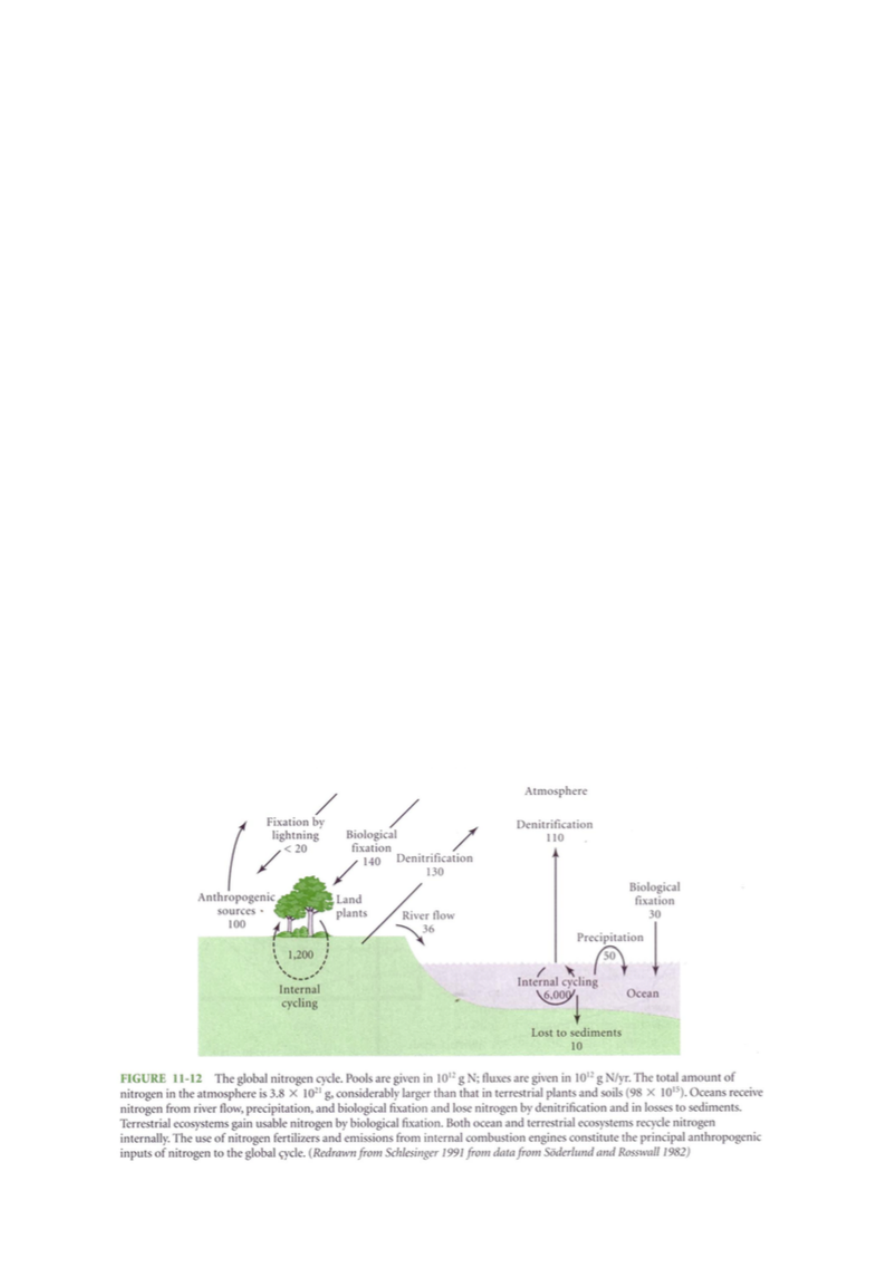




Kõik kommentaarid