Ökoloogia kordamisküsimused 2022 (0)
Elu - Luuletused, mis räägivad elus olemisest, kuid ka elust pärast surma ja enne sündi.
Ökoloogia eksami (LTOM 01.105) kordamisküsimused 2022
1. PÕHIMÕISTED
Kooslus – kooseksisteerivate organismide kogum.
Ökoloogia – teadus organismide ja nende keskonna vahelistest suhetest (vastasmõjudest,
interaktsioonidest (Ernst Haeckel) / teadus eluslooduse struktuurist ja süsteemist (Eugene
Odum).
Biotsönoos – elusorganismide kooslus.
Ökosüsteem – mingi elupaiga elukooslust pluss viimase elutut keskkonda (Arthur Tansley).
Biogeotsönoos – looduslik kompleks, millesse kuuluvad elukooslus (biotsönoos) ja selle
elupaiga (biotoobi, ökotoobi) eluta keskkond.
Kohastumine ehk fülogeneetiline adaptatsioon – genoomi muutustega seotud keskkonnaga
sobitumine.
Kohanemine ehk ontogeneetiline adaptatsioon – organismi eluea jooksul ilma genoomi
muutusteta toimuv keskkonnaga sobitumine.
Unitaarsed organismid – organismid on ajalis-ruumiliselt ja geneetiliselt selgelt eristuvad.
Modulaarsed organismid – organismid, mis koosnevad vähem või rohkem eristatavatest ja
vähem või rohkem iseseisvatest moodulitest, mis on geneetiliselt identsed.
Klonaalne paljunemine – paljunemine, mille puhul järglased ei erine emast geneetiliselt
(taimede puhul nimetatakse vegetatiivseks paljunemiseks).
Risoom – taimede harunev või harunemata, horisontaalse, vertikaalse või tõusva suunaga,
mitmeaastane, maa-alune võsu.
Stoloon – on varre maapealne pikenenud külgharu, millel tekkivate pungade abil taim levib
ning vegetatiivselt paljuneb.
Apomiksia – on suguta paljunemine, kus seeme areneb viljastamata munarakust ja sellest
idanev järglane on emataime ”identne kaksik”.
Partenogenees – on suguta paljunemine, kus embrüo areneb viljastamata munarakust
(loomade puhul).
Võsu ehk ramet – iseseisva või potentsiaalselt iseseisva juuresüsteemiga taimemoodul,
olenemata sellest kas tegemist on klonaalselt või seemnelist päritolu mooduliga (taimede
puhul).
Populatsioon – ühe ja sama liigi esindajate piiritletavat kogumit, (mis koosneb omavahelt
potentsiaalselt vabalt ristuvatatest isenditest) / mingi liigi mingil ajal mingis kohas.
Panmiktiline populatsioon ehk deem – vabalt ristuvate isendite kogum, pole
populatsioonisiseseid paljunemisbarjääre.
Levis ehk diaspoor – on igasugune organismide (eriti taimede ja seente) levimisalge, ka
organismi osa, mille abil organismid levivad (eosed ehk spoorid, seemned).
Bioom ehk makroökosüsteem – sarnastes klimaatilistes ja ökoloogilistes tingimustes levivad
ning sarnase struktuuriga ökosüsteemid üle maailma.
Ökofüsioloogia – ökoloogia piirteadus, mis uurib ökoloogilisi protsesse organelli ja organi
tasemel.
Autökoloogia – ökoloogia valdkond, mis tegeleb uuringutega isendi tasemel.
Demökoloogia – ökoloogia valdkond, mis tegeleb uuringutega populatsiooni tasemel.
Sünökoloogia – ökoloogia valdkond, mis tegeleb uuringutega koosluse tasemel.
Süsteemökoloogia – ökoloogia valdkond, mis tegeleb uuringutega kogu ökosüsteemi
tasemel.
2. AUTÖKOLOOGIA
Ressursifaktorid ehk ressursid – energia või aine vood, mida organismid otseselt
assimileerivad, s.t. oma kehasse võtavad ja enda osaks muudavad.
FOTOSÜNTEETILISELT AKTIIVNE KIIRGUS (PAR)
Fotosünteetiliselt on aktiivne selline elektromagnetkiirgus, mis jääb vahemikku 380-710 nm.
Päikeselt saabuvast kiirguse koguhulgast moodustab PAR ca 44%. PAR-ist lühema
lainepikkusega on ultraviolettkiirgus (UV), PAR-ist pikema lainepikkusega on kaugpunane
ehk infrapunane kiirgus (FR; nimetatakse ka soojuskiirguseks). Enam-vähem täpselt
samasse vahemikku kui PAR jääb ka kiirgus, mida inimese silm tajub valgusena.
Fotosünteesiaparaadis on keskne roll klorofüllil, mis suudab PAR-i
lainepikkusega ergastuda ja konverteerida neelatud footoni energia
keemiliseks energiaks. Klorofülli on looduses kahte tüüpi: klorofüll
a ja klorofüll b. Kõige efektiivsemalt assimileerivad
klorofüllimolekulid sinist ja punast kiirgust, seetõttu näivad ka
taimelehed rohelised. Kloroplastides sisalduvad lisapigmendid –
karotinoidid ja ksantofüllid, mis suudavad ka rohelisest kiirgust
assimileerida.
Fotosünteesi efektiivsus – proportsioon peale langevast PAR-ist,
mis keemiliseks energiaks muundatakse. Sõltub: peegeldumiskoefitsient (taimelehtede pinda
katva ja kaitsva vahakihi ehk kutiikula paksus); läbivuskoefitsient (taimelehe paksus, kihid ja
plastiidide paiknemine lehes; tõenäosus, et lehes neeldunud footon tabab klorofüllimolekuli
(või lisapigmendi molekuli) ja ergastab selle. Looduslikes ökosüsteemides on alla 0,03 (st
3%). Troopikas Ef = 2-3%, parasvöötmes Ef = 0,6-1,2%.
Kihilisust mõõdetakse maismaaökosüsteemides koosluse lehepinnaindeksi abil (LAI). LAI
on dimensioonitu koefitsient, mis mõõdab, kui palju lehepindala tuleb ühe ühiku maapinna
kohta. LAI on suurim ekvatoriaalsetes vihmametsades ja väikseim kõrbetes.
Taimekoosluste rindelisus (lehestiku jagunemine eristatavateks ja tihti ka erinevatest
liikidest koosnevateks kihtideks) tähendab automaatselt seda, et taimede hulgas peab leiduma
nii valgus- kui ka varjutaimed. Valgus- ja varjutaimedel on erinev fotosünteesi
kompensatsioonipunkt – liigiomane PAR-i minimaalne tase, mis tagab taime nullkasvu
(taim on elus kuid ei kasva).
Fotosünteesikõverad on kõverad, kus kujutatakse fotosünteesi intensiivsuse sõltuvust PAR-i
kättesaadavusest. Kõrge kiirgustase põhjustab varjuliigidel fotoinhibitsiooni – fotosüntees
läheb „lukku“ ja taime kasv aeglustub või lakkab.
SÜSINIK
Autotroofid tarbivad süsinikuühendeid peamiselt
mineraalsel kujul. Maismaa primaarprodutsentidele on
mineraalse süsiniku allikana süsihappegaas (CO2).
Veekeskkonnas on autotroofid võimelised tarbima
süsinikku lisaks vees lahustunud süsihappegaasile ka
süsihappena
(H2CO3) või ioonsel kujul –
vesinikkarbonaatioonina (HCO3-) või karbonaatioonina (CO32-). Sealjuures sõltub
veekogudes erinevat sorti mineraalsete süsinikuallikate kättesaadavus otseselt vee
reaktsioonist (pH). Kõrge pH-ga (aluselistes) vetes assimileeritakse peamiselt
karbonaatioone, happelistes vetes peamiselt CO2 ja süsihapet.
Tänapäeval on süsihappegaasi sisaldus maakera
atmosfääris ca 400 ppm ehk 0.04%. See on
evolutsioonilises ajaskaalas rekordiliselt madal number.
Karboni ajastu lõpul, ca 280 miljonit aastat tagasi oli
CO2 kontsentratsioon atmosfääris võrreldavalt madal.
Hulkrakse elu hoogsa evolutsioneerumise perioodil
(„kambriumi plahvatus“, 500+ miljonit aastat tagasi) oli
CO2 sisaldus atmosfääris praegusega võrreldes 14-18
korda kõrgem, kriidi ajastu keskel (ca 100 miljonit
aastat enne meid) oli see umbes kuus korda suurem kui
praegu. Viimastel aastakümnetel on tõus olnud enam-vähem lineaarne ja isegi väikese
kiirendusega, atmosfääri süsinikuvaru on tänapäeval ligi 30% suurem kui see oli 60 aastat
tagasi.
Kasvuhooneefekti tekkimiseks peab gaasimolekul olema ebasümmeetrilise ehitusega. Klaas
või läbipaistev plastik laseb läbi nähtavat valgust, kuid takistab kaugpunakiirgusel tagasi
maailmaruumi püüdlemist, ja osa kiirguse energiast jääb kasvuhoonesse lõksu, soojendades
kasvuhoone sisekeskkonda kõrgema temperatuurini kui väljas. Kasvuhoonegaasid: CO2,
veeaur, metaan, lämmastikoksiidid, osoon, aga mitte kahest aatomist gaas (O2, H2, N2) ja
väärisgaas.
Fotosünteesi (ja vastupidise protsessi – aeroobse rakuhingamise ehk oksüdatiivse
respiratsiooni) üldvalem on: 6CO2 + 6H2O + e ↔ C6H12O6 + 6O2.
Vähese fotosünteesi efektiivsuse tingib valgushingamine ehk fotorespiratsioon Calvini
tsüklis süsiniku esmase fikseerimise eest vastutava ensüümi RuBisCo tõttu. RuBisCo on
võimeline süsinikdioksiidi asemel siduma hoopis molekulaarset hapnikku, raisates sellega
energiat ning süsinikku.
Evolutsioonis on fotorespiratsiooni kahjulike mõjude leevendamiseks välja arenenud kaks
energeetiliselt kokkuhoidlikku fotosünteesitüüpi – C4- ja CAM-fotosüntees (C4- ja CAM-
taimed). Need vastanduvad C3-fotosünteesile, mis on omane enamikule maakera
taimeliikidest (C3-taimed).
C3-taimedel pole spetsiaalseid kohastumusi fotorespiratsiooni pidurdamiseks. Calvini tsükli
esimeseks etapiks on CO2 sidumine RuBisCo külge, tulemuseks on kolme süsinikuaatomiga
süsinikuühend.
C4-taimedes on PAR-ist sõltuvad keemilised reaktsioonid ja Calvini tsükkel üksteisest
füüsiliselt eraldatud. Valgusreaktsioonid toimuvad mesofüllirakkudes, Calvini tsükkel leiab
aset kimbuümbrise rakkudes. Mesofüllirakus fikseeritakse süsinik nelja süsinikuaatomiga
orgaanilisse happesse nimega oksaloatsetaat. Seda protsessi vahendab ensüüm PEP-
karboksülaas, millel ei esine tendentsi molekulaarset hapnikku siduda. Oksaloatsetaat
konverteeritakse järgnevalt malaadiks, mida on võimalik kimbuümbrise rakkudesse
transportida. Viimastes malaat laguneb, vabastades CO2 molekuli. Seejärel seotakse CO2
RuBisCo poolt ja see siseneb Calvini tsüklisse.
CAM (paksulehelise happe-metabolism) -taimed hoiavad lehtede õhulõhed avatuna ainult
öösiti, võimaldades atmosfäärsel CO2-l lehtedesse difundeeruda ja kaotades samaaegselt
jaheda välistemperatuuri tõttu väheses koguses vett. Süsinik fikseeritakse seejärel
oksloatsetaadina PEP-karboksülaasi abil. Seejärel konverteeritakse oksaloatsetaat malaadiks,
mida säilitatakse rakkude vakuoolides kuni järgmise päevani. Päevase intensiivse
valguskiirguse käes CAM-taimed õhulõhesid üldse ei ava, kuid suudavad siiski
fotosünteesida. See on võimalik, kuna orgaaniline hape transporditakse vakuoolist välja ja
lõhustatakse, vabastamaks CO2, mis seejärel siseneb Calvini tsüklisse.
HAPNIK
Molekulaarse hapniku sisaldus tänapäevases atmosfääris on 20,1%. Fotosünteesi käigus
vabaneb iga redutseeritud süsinikuaatomi kohta üks hapnikumolekul, mis hingamise käigus
uuesti süsiniku külge seotakse. Vabanenud hapnikuaatomid ei pärine mitte CO2 küljest,
vaid veemolekulist – päikeseenergia toel toimub fotosünteesi käigus vee fotolüüs, s.t. hapnik
kistakse vesiniku küljest lahti ja heidetakse atmosfääri või veekeskkonda.
Atmosfääri hapnikusisalduse dünaamikas on eristatavad viis staadiumit:
Staadium 1 (3,85-2,45 mld aastat tagasi) –
hapnikku atmosfääris praktiliselt pole;
Staadium 2 (2,45-1,85 mld aastat tagasi) –
O2 toodetakse sinivetikate poolt, kuid
enamik sellest jääb ookeani ning seotakse
ookeanipõhja kivimite poolt;
Staadium 3 (1,85-0,85 mld aastat tagasi) –
O2 jõuab atmosfääri, kuid kulub peamiselt maismaakivimite oksüdeerimiseks ning
osoonikihi moodustumiseks stratosfääri;
Staadium 4 (0,85-0,57 mld aastat tagasi) – hapniku keemiline sidumine kivimitesse
on lõppenud ning O2 hakkab kiiresti atmosfääri kogunema;
Staadium 5 (0,57-0 mld aastat tagasi) – pärast mitmeid võnkeid jääb atmosfääri
hapnikusisaldus 20% kanti pidama.
Hapnik võib limiteerida sekundaarproduktsiooni. Maismaaökosüsteemides (soodes)
juhtub see liigniisketes ja seisva või väheliikuva põhjavee tingimustes, kus mullavesi rikastub
molekulaarse hapnikuga väga aeglaselt. Heterotroofide rakuhingamine kulutab mullast ära
kogu vees lahustunud O2 ja aeroobne heterotroofne produktsioon aeglustub või lakkab.
Veeökosüsteemides toimub fotosüntees ainult pinnakihis, sest sügavamal kui 200 m on
enamik PAR-ist vees neeldunud ja keskkond liiga pime. Hapnikupuudusel jääb ka
süvaookeanis (nagu soodes) osa orgaanilistest süsinikuühenditest lagundamata ning
akumuleerub põhjasetetena.
Sekundaarproduktsiooni limiteerib hapnik soojades vetes sagedamini kui külmades, kus
hapniku lahustuvus on parem. Temperatuuri tõustes 0o kuni 35o C-ni väheneb O2 lahustuvus
ca kaks korda. Hapniku lahustuvus paraneb oluliselt ka sügavusega.
Kõige sagedamini näeme hapnikupuudust ja sellest tulenevat heterotroofse elu kidumist või
hukkumist külmas kliimas paiknevates madalates ja paljutoitelistes (eutroofsetes) ning vee
aktiivse läbivooluta mageveekogudes, mis talvel jäävad paksu jääkaane alla. Vähese
mineraalainete sisaldusega vähetoitelistes ehk oligotroofsetes siseveekogudes hapnikunälga
üldjuhul ei teki.
MINERAALSED ELEMENDID
Elemendid, mis on vajalikud kõikidele organismidele ja mõnele kindlale organismirühmale.
Vastavalt elutegevuseks vajalikule kogusele jagatakse toiteelemente makro- ja
mikroelementideks. Makroelemente jaotatakse omakorda esmasteks ja teisesteks, sõltuvalt
vajaminevast kogusest. Esimeste hulka kuuluvad lisaks struktuursetele elementidele (C, O,
H) veel N, P ja K. Teisesteks makroelementideks on Ca, S ja Mg.
Lämmastik N kuulub iga aminorühma (-NH3) koosseisu ja on seetõttu hädavajalik valkude
sünteesimiseks. Taimed assimileerivad lämmastikku kas nitraatioonina (NO3-),
nitritioonina (NO2-) või ammooniumioonina (NH4+). Lämmastikku sisaldavad soolad on
hästi dissotsieeruvad ning seetõttu lämmastikuühendid väga liikuvad ökosüsteemis ja
leostuvad mullast kergesti välja.
Fosfor P kuulub oluliste biomolekulide koostisse (nukleiinhapped, ATP) ja osutub
kriitiliseks ehk taimekasvu limiteerivaks elemendiks. Taimed assimileerivad fosforit mullast
peamiselt divesinikfosfaat- (H2PO4-) või vesinikfosfaatiooni (HPO42-) kujul.
Kaalium K ei kuulu olulise keemilise ühendi koosseisu. K osaleb süsivesikute ja valkude
sünteesis, taimede sisekeskkonna veesisalduse regulatsioonis ning katalüüsib mitmete
oluliste makromolekulide sünteesi.
Väävel S on eluks hädavajalik kõigi taimede kasvuks ja füsioloogiliseks toimimiseks
(tsüsteiin ja metioniin). Mullast assimileerivad taimed väävlit sulfaatidena kuid see tuleb
taimedes enne metabolismi lülitamist redutseerida sulfiidideks.
Kaltsium Ca, mida assimileeritakse katioonina (Ca2+), on taimedele oluline kui teiste
mineraalsete toitainete assimileerimise regulaator ning osaleb kaltsium taimeensüümide
aktiveerimisel.
Magneesium Mg on rohelistele taimedele hädavajalik, sest iga klorofüllimolekuli kuulub üks
magneesiumiaatom.
RESSURSSIDE SÜNERGEETILINE MÕJU ORGANISMIDELE
Sünergiaks ehk samasuunaliseks koostoimeks nimetatakse tavaliselt faktorite summaarse
efekti suurenemist mitme üheaegse teguri koostoimel.
Kaks ressurssi üksteise suhtes võivad olla kas:
Hädavajalikud (essentsiaalsed) – peab organism parema elutegevuse intensiivsuse
nimel tarbima mõlemat ressurssi sest ühte teisega asendada ei saa (näiteks PAR ja
mineraalsoolad taimedel);
Asendatavad – üks ressurss võib teist ka täiel määral asendada (näiteks kodukanade
toidulaual võib nisu asendada odraga);
Nimetatud kahel juhul ressursside vahel sünergia puudub.
Komplementaarsed – ressursse koos tarbides on summaarne kulu väiksem kui ainult
üht neist assimileerides (näiteks rohusööjad tarbivad üht ja sama taimeliiki, võivad
seal leiduvad toksiinid organismis kuhjuda ja tervise ära rikkuda);
Anatagonistlikud – kaht ressurssi läbisegi tarvitades summaarne ressursivajadus
suureneb (näiteks niiskestroopikas, kus tolmeldajate meelitamiseks mõeldud ressursid
eraldi võetuna toksilised ei ole kuid erinevate taimeliikide toksiinid koos tarvitades
moodustavad „mürgise kokteili“).
Tingimusfaktorid ehk tingimused kas võimaldavad, või siis ei võimalda, ressursse
elutegevuse jaoks tarbida.
TEMPERATUUR
Mida tugevam on ookeani mõju maismaa kliimale, seda väiksem on temperatuuri sesoonne
varieerumine (seda suurem merelisus ja seda väiksem kontinentaalsus).
Enamik teadaolevaid organisme on kõigusoojased
(poikilotermsed, külmaverelised) – nende keha
sisetemperatuur sõltub peamiselt väliskeskkonna
temperatuurist. Ainult linnud ja imetajad on püsisoojased
(homöotermsed, soojaverelised), s.t. neil eksisteerivad
füsioloogilised mehhanismid konstantse ja enamasti
välistemperatuurist kõrgema (aga vajadusel ka madalama)
sisetemperatuuri püsivaks hoidmiseks.
Sesoonses kliimas elavatel kõigusoojastel, näiteks taimedel, saab kindlate ontogeneesi
järkude (fenofaaside) ajastust üsna täpselt ennustada aktiivsete temperatuuride summaga.
Aktiivseteks loetakse taimede puhul selliseid ööpäeva keskmisi
temperatuure, mis ületavad 5 oC; näiteks kaks ööpäeva
keskmise temperatuuriga +20 oC annab kolmkümmend
kraadpäeva (2 päeva * (20 – 5 oC) = 30).
Maismaaökosüsteemides on aasta keskmine temperatuur
keskse tähtsusega tingimusfaktor, mis kombineerituna aasta
keskmise sademete hulgaga lubab edukalt kahemõõtmeliselt
ordineerida (üksteise suhtes järjestada või paigutada) maakeral
levivaid bioome.
KESKKONNA pH
pH on keskkonna happesuse mõõt, defineeritud kui
negatiivne kümnendlogaritm aktiivsete H+- ioonide
kontsentratsioonist mulla lahuses. Mulla võib lahuse
reaktsioon olla kas neutraalne (pH = 7), happeline (pH < 7)
või leeliseline (pH > 7). Mulla pH mõjutab taimedele
vajalike mineraalelementide kättesaadavust ning määrab,
millises keemilises vormis need mullas esinevad ja millistes keemilistes reaktsioonides
osalevad.
Happelised mullad on valdavalt levinud suure
sademete hulgaga piirkondades – vihmavesi on
nõrgalt happelise reaktsiooniga (pH ≈ 5,7). Mulla
happesust võib tõsta: mitmete maapõues leiduvate
mineraalide
oksüdatiivne porsumine
ehk
murenemine keemiliste protsesside tagajärjel;
taimede
juurhingamine, juurtoitumine
ja
orgaanilise aine lagundamine.
Aluseliste muldade tekke peamiseks eelduseks on
mulla lähtekivimi, s.t kivimi, mille murenemisel muld tekib. Aluseliste ühendite
akumulatsioon mullas toimub juhul, kui läbi mulla ei liigu piisavalt vett, et vees lahustuvaid
soolasid mullast välja leostada.
SOOLSUS
Soolsuseks nimetatakse vees lahustunud või mullas leiduvate mineraalsoolade
kontsentratsiooni. Vett soolsusega alla 0,5‰ nimetatakse mageveeks. Kui soolsus jääb
vahemikku 0,5-30‰, on tegemist riimveega (laguunid, sood, järved ja ookeanidega kitsaste
väinade kaudu ühendatud mered). Üle 30‰ soolsusega vett kutsutakse soolaseks veeks.
INFORMATSIOON
Kemoretseptsiooni on võime ümbritseva keskkonna keemilisi omadusi tajuma ning neile
adekvaatselt reageerima, selleks on võimelised kõik organismid (maitse ja lõhna tundmine).
Stressiseisundis naabertaimede poolt eritatavad kergesti lenduvad madalmolekulaarsed (nn
volatiilsed) keemilised ühendid on paljudele taimeliikidele hindamatu informatsiooni
allikaks.
Fotoretseptsiooni ehk elektromagnetkiirguse taju suudavad kõik fotosünteesivad organismid,
aga ka suur enamus loomadest. Liike, kes tajuvad sellise lainepikkusega kiirgust, mis täiesti
või valdavalt kattub PARiga. Aga ka UV või FR kiirguse retseptsiooni võime on üsna
levinud, vastavalt näiteks putukatel või roomajatel.
Mehhanoretseptsioon ehk taktiilse retseptsiooni võime. Organismid tajuvad mehhaanilist
survet, tihti ka neid ümbritsevate keskkonnagaaside või -vedelike liikumist (näiteks on
kaladel olemas küljejooneelund, mis varustab isendit pidevalt infoga vee liikumise kiiruse
ning suuna kohta; mullast või veest loomset toitu otsivatel lindudel on võime nokas
paiknevate Herbsti kehakeste abil tunnetada vibratsioone, mida liikumisel tekitavad putukad
või väikesed kalad).
Audioretseptsioon ehk võime helisid tajuda. See on omane ainult lülijalgsetele ja
selgroogsetele. Kajalokatsioon, mille käigus loomad väljastavad ultrahelisignaale ja
suudavad tajuda helilainetele ette jäävatelt objektidelt tagasi peegeldunud signaale
(nahkhiired, delfiinid, vaalad).
Baroretseptsioon on veekeskkonnas elutsevatel loomadel võime keskkonna rõhku tajuda ja
saadud info järgi oma füsioloogiat vastavalt sügavusele kohandada. Nad kasutavad, et
spetsiaalseid elundeid kasutades keha erikaalu muuta ning soovitud sügavusel püsida (kaladel
on ujupõis).
Magnetoretseptsioon on võime tajuda maa magnetvälja suunda ja selle järgi ruumis
orienteeruda. Organismide sisemised kompassid on väga tundlikud ja peamist rolli mängivad
seal kergesti magnetiseeruvat rauaühendit – magnetiiti – sisaldavad koed või organid.
Magnetiit on ferrimagneetiline aine. Lindudel võib magnetiiti leida silma võrkkestas ehk
reetinas.
Elektroretseptsioon on võime tajuda teiste organismide tekitatud elektrivälja (haidel, raidel
ja kuuel imetajaliigil). Kasutavad nimetatud loomad magnetilist induktsiooni (magnetvälja
mõju iseloomustavat füüsikalist suurust, mis väljendab väljas liikuvale elektrilaengule
mõjuvat jõudu). Sensoorseid elundeid kutsutakse Lorenzi ampullideks.
NIŠITEOORIA
Vaadeldava ökoloogilise faktori väärtuste vahemikku, mille puhul liik kasvada ja paljuneda
suudab, nimetatakse liigi ökoloogiliseks amplituudiks.
Lisaks faktori miinimumile ja maksimumile on igal liigil
enamike faktorite suhtes välja kujunenud ka optimum
ehk faktori väärtus, mille puhul organismi elutegevuse
intensiivsus on maksimaalne. Optimumi olemasolul
illustreerib elutegevuse intensiivsuse ning faktori väärtuse
vahelist seost tolerantsikõver.
Tolerantsikõver võib olla ekvivalentne normaaljaotuse tihedusfunktsiooni graafikuga, s.t. on
kellukesekujuline ja optimumi suhtes peegelsümmeetriline, aga võib olla mitte
peegelsümmeetriline. Keskmine graafik kujutab tolerantsikõverat, siis kui faktoriks on
toksiliste ainete sisaldus keskkonnas (saasteained jms). Organismid neid aineid ei vaja, vaid
kannatavad nende olemasolu kuni teatava piirini ja siis kipuvad ära surema. Kuna mürke ei
vajata, siis puudub organismil nende jaoks ka optimum.
Tolerantsikõveral on kolm parameetrit:
Tolerants ehk ökoloogilise amplituudi laius (ulatus). Kitsalt mingile faktori
väärtusele spetsialiseerunud liikidel, nn spetsialistidel ehk stenotoopidel on tolerants
väike; laiale faktori vahemikule kohastunud liikidel – generalistidel ehk
eurütoopidel – on tolerants suur;
Optimum;
Maksimaalne elutegevuse intensiivsus, mille organism optimaalsetel tingimustel
saavutab.
Ökoloogiline amplituud on käsitletav ka kui organismi ühemõõtmeline ökoloogiline nišš.
Nišš on piirkond n-mõõtmelises faktorruumis, millele liik on kohastunud.
Ühemõõtmeline nišš on tegelikult sama, mis
ökoloogiline amplituud. Vaadeldakse üht olulist
keskkonnafaktorit korraga. Kahemõõtmelise niši
korral on vaatluse kaks faktorit korraga.
Kolmemõõtmelise niširuumi korral on vaatluse
kolm faktorit korraga.
Keskkonnatingimusi, milles vaadeldav liik oleks ilma naaberorganismide piirava mõjuta
teoreetiliselt võimeline püsivalt elutsema, nimetatakse
potentsiaalseks
ehk
autökoloogiliseks nišiks. Tegelik ehk realiseerunud (sünökoloogiline) nišš on alati
väiksem kui potentsiaalne, kuna teatavatest potentsiaalse niši piirkondadest on liik teiste
organismide poolt välja tõrjutud.
Liebigi miinimumi reegel: organismide kasvu ning paljunemist limiteerib see ressurss, mille
tase on ökoloogilisele miinimumile suhteliselt kõige lähemal.
Shelfordi tolerantsi reegel: organismide kasvu ja paljunemist limiteerib see ressurss, mille
tase on optimumist suhteliselt kõige kaugemal.
FENOTÜÜBILINE PLASTILISUS
Organismide plastilist vastust keskkonnale võib võib jagada kaheks põhimõtteliselt erinevaks
komponendiks – paratamatu ehk passiivne plastilisus ja adaptiivne ehk aktiivne
plastilisus.
Passiivne plastilisus tuleneb tõigast, et paljud organismi fenotüübilased tunnused on otseses
seoses isendi kehasuurusega. Selliseid positiivseid sõltuvusi organismide mitme erineva
tunnuse vahel nimetatakse allomeetrilisteks seosteks, reeglina tähendab see lineaarset seost
logaritmitud kehamassi ja logaritmitud tunnuse väärtuse vahel.
Aktiivne plastilisus on alati seotud assimileeritud ressursside erineva ümberpaigutamise ehk
allokatsiooniga erinevate funktsioonide tarbeks. Aktiivne plastilisus avaldub lõivsuhtena –
negatiivse seosena organismi erinevate võimekuste vahel.
Allomeetrilise seose
tekitab üldjuhul populatsioonisisene varieeruvus üldise
ressursihõivamise võimekuses, lõivsuhte tekitab varieeruvus omastatud ressursside jagamises
erinevate funktsioonide vahel.
3. DEMÖKOLOOGIA
ÜKSIKU POPULATSIOONI KASV
Populatsiooni ajalise dünaamika vaatlemisel on keskse tähtsusega populatsiooni liikmete arvu
ehk populatsioonitiheduse (N) muutus ajas (t) ja samuti muutumise suund, mida väljendab
populatsiooni kasvukiirus dN/dt. Kui populatsioon ajas suureneb, on kiirus positiivne,
vähenedes negatiivne. Nullkasv (dN/dt = 0) tähendab populatsiooni liikmete arvu püsimist
samal tasemel.
Populatsiooni piiramatu kasvu võrrand ehk populatsiooni eksponentsiaalse kasvu võrrand
ehk Malthuse võrrand: dN/dt = bN – dN, kus b – liigiomane sünnikoefitsient ehk -
parameetr, d - liigiomane surmakoefitsient ehk -parameetr.
dN/dt = rN, kus r (alati positiivne arv) on erikasvukiirus ehk sisemine kasvukiirus ehk
biootiline potentsiaal, mis näitab liigi võimet ajaühikus ühe populatsiooniliikme kohta
järglasi toota. Võrrand illustreerib, et populatsiooni kasv on autokatalüütiline ehk
positiivselt tagasisidestatud protsess – „mida rohkem, seda rohkem; mida vähem, seda
vähem“ (negatiivne tagasiside tähendab regulatsiooni: „mida rohkem, seda vähem; mida
vähem, seda rohkem“.
Ressursse on ökosüsteemis alati piiratud hulgal ja mingi populatsioonitiheduse juures hakkab
ressursipuudus kasvu piirama, tekib keskkonnatakistus (R), R = aN2, kus a on liigi- ja
keskkonnaomane parameter.
Populatsiooni piiratud kasvu võrrand ehk logistilise kasvu võrrand: dN/dt = rN(1 – N/K) =
rN((K – N)/K), kus K = r/a on keskkonna kandevõime. Keskkonna kandevõime (K)
näitab, kui suurt vaadeldava liigi populatsiooni on keskkond võimeline kandma, ehk siis
majutama ja ära toitma. Populatsiooni kasvukiirus on positiivne, kui N < K, N = K on
populatsioon nullkasvus, kui N > K on kasv negatiivne.
Piiratud kasvu võrrandis on kaks muutujat (N, t) ja kaks liigiomast parameetrit (r, K). Kui
r on kõrge, siis liik on kiire paljuneja, kui K on kõrge, siis liik kasutab ressursse efektiivselt ja
ökosüsteemis on korraga võimelised elatuma ning paljunema suur hulk isendeid.
DEMOGRAAFILISED PARAMEETRID
Dünaamiline meetod ehk kohordi meetod, kui populatsiooniprotsesse vaadeldakse reaalajas,
jälgides ühe põlvkonna ehk kohordi saatust sünnist surmani. Sobib selline lähenemine ainult
lühikese elueaga liikide uurimiseks.
Staatiline meetod ehk populatsiooni demograafilise püramiidi meetod, kui tehakse
minevikus ja olevikus kulgevate protsesside kohta järeldusi populatsiooni vanuselise
struktuuri järgi mingil ajahetkel.
Demograafilised parameetrid:
lx – ellujäämus ehk elumus: tõenäosus saavutada vanus x, proportsioon algsest
kohordist (x=0), kes elab vanuseni x;
dx – suremus: tõenäosus surra vanuses x, proportsioon algsest kohordist, kes sureb
vanuses x;
qx – vanusest sõltuv surmarisk: x-vanuses kohordi liikme tõenäosus surra ilma
vanust x+1 saavutamata, proportsioon x-vanuse saavutanud isenditest, kes sureb sama
staadiumi jooksul;
mx – viljakus: keskmine järglaste arv ühe vanuses x kohordi liikme kohta selle
vanusejärgu jooksul;
Fx – kohordi summaarne viljakus: kohordi liikmete toodetud järglaste koguarv
vanuses x.
R0 – populatsiooni puhas kasvukiirus: järglaste arv, mida üks kohordi liige
keskmiselt elu jooksul toodab.
, kus T tähistab generatsiooni kestust ehk ühe kohordi maksimaalset eluiga.
Kui R0 > 1 siis populatsioon kasvab, kui R0 = 1 siis populatsiooni suurus on
ajas konstantne, kui R0 < 1 siis populatsioon kahaneb
Semelpaarne (monokarpne) liik, ehk selline, kes teeb elu jooksul ainult ühe
paljunemispingutuse (viljakad on ainult kõrgeima vanuseklassi esindajad).
Iteropaarne (polükarpne) liik, ehk selline, kes on elu jooksul suutelised korduvalt
paljunema.
Paljunemisväärtus (vx) ehk ühe isendi keskmine tulevase põlvkonna ootus
mingis vanuses (x). Valemis tähistab (t) eluiga alates käesolevast vanusest
(x) kuni viljaka eluea lõpuni (T).
Deevey kõverad ehk elumuse kõverad, mis näitavad, kuidas on logaritmitud elumuse (ln lx)
paratamatu sünnijärgne monotoonne langus seotud isendi vanusega.
Esimest tüüpi kõverad iseloomustavad organisme, kelle
puhul suremus koondub peamiselt vanematesse
elujärkudesse (qx ehk vanusest sõltuv surmarisk kasvab
monotoonselt elu jooksul). Teist tüüpi kõver tähistab
teoreetilist olukorda, kus qx on elu jooksul konstantne (st
vanusest sõltumatu). Kolmandat tüüpi kõverad
iseloomustavad organisme, kelle puhul suremus on
peamiselt koondunud varastesse elujärkudesse (qx langeb
elu jooksul monotoonselt), suguküpseks saab vähe isendeid, kuid need elavad kaua.
Demograafiliseks püramiidiks nimetatakse graafikut, kus X-telg
mõõdab mingi vanusega isendite osakaalu populatsioonis, Y-telg aga
populatsiooniliikme vanust. Traditsiooniliselt kujutatakse
demograafilist püramiidi peegelsümmeetrilisena Y-telje suhtes nii, et
isaste isendite arv on märgitud vasakule poolele ja emaste arv
paremale poolele Y-telge. Noore ja kasvava populatsiooni
demograafiline püramiid on laia põhjaga ja kitsa tipuga. Vananeva
ja taanduva populatsiooni demograafiline püramiid on „raske“ tipu
ja kitsa alusega. Stabiilse suurusega jätkusuutliku populatsiooni püramiid jääb kujult kahe
eelmise vahele.
LEVIK, LEVIMINE, MIGRATSIOON
Levik on isendite paiknemine ruumis, asetumine ruumis üksteise suhtes, jaotus ruumis.
Levimine on protsess, mis leviku taga seisab, liikumine ruumis. Juhtumit, kus mingi
bioloogiline liik omandab võime lühikese aja jooksul edukalt ja massiliselt uutesse
ökosüsteemidesse levida ning seal tähelepanuväärse osa ressurssidest omastada sellega
eluskoosluse senist toimimist ümber korraldades, nimetatakse invasiooniks ja edukat liiki
invasiivseks liigiks ehk invandiks. Migratsioon ehk ränne – suure hulga isendite suunatud
ja populatsiooniökoloogiliselt mittelõplik liikumine ühest paigast teise.
Ruumilise jaotuse tüübid:
Juhusliku ruumilise jaotuse korral paiknevad isendid
ruumis üksteisest sõltumatult. Näide: kauglevivad
üheaastased seemnetaimed, vees passiivselt hõljuvad
organismid.
Regulaarse ruumilise jaotuse korral isendid püüavad
miskipärast omavahelist distantsi maksimeerida. Näide:
linnuliikidel pesa asetseb enamvähem territooriumi keskel, laialdase juurestikuga
mitmeaastastel kõrbetaimedel asub iga maapealne võsu suure “toitumisterritooriumi” keskel.
Agregeeritud ruumilise jaotuse korral isendid mingil põhjustel koonduvad salkadesse.
Näide: kogunemine toiduobjekti lähedusse (näiteks kahjurid teatud liiki puu võras), klonaalse
levimise puhul koonduvad ühe organismi järglased emaorganismi lähedusse, paljud
loomaliigid koonduvad karjadesse, parvedesse või peredesse.
Levimine võib olla aktiivne (liikumisvõimelistel organismidel) ja passiivne (näiteks
tuullevijatel taimedel, vooluvetega levivatel liikidel). Lisaks on paljudel taimeliikidel olemas
paiskviljad – seeme heidetakse valmimisel mehhaaniliselt emataimest eemale. Sessiilsed
organismid on loomad, kes on vanemate elujärkude jooksul liikumatud. Neil on mingil
varasel eluperioodil olemas aktiivselt või passiivselt liikuvad diaspoorid.
Migratsiooni tüübid:
Korduvkasutusega pilet. Migreerutakse kord aastas toitumispaika ja kord aastas
paljunemispaika (sesoonne migratsioon). Näide: iteropaarsed liigid: rändlinnud,
siirdekalad, pingviinid.
Edasi-tagasi pilet. Migreerutakse üks kord elus toitumispaika ja üks kord elus
paljunemispaika. Näide: semelpaarsed liigid: lõhelised (toituvad ookeanis, paljunema
lähevad elu lõpul jõgedele), angerjas (toitub üleilmselt, paljuneb ainul Sargasso
meres), paljud sääsed (munevad vette, toituvad maismaal).
Üheotsa pilet. Kord elus migreerutakse ühest paigast teise ja paljunetakse seal,
järgmine põlvkond migreerub tagasi lähtekohta. Näide: semelpaarsed liigid: admiral
(Suurbritannia lõunaosast migreeruvad Ibeeria poolsaarele. Seal munetakse ja
surrakse. Järgmine põlvkond migreerub Suurbritanniasse) ja monarhliblikad.
LIIGISISENE KONKURENTS
Taimekooslustes toimub konkurentsivõitlus korraga nii mullas (juurkonkurents) kui
taimkatte võras (võsukonkurents ehk valguskonkurents).
Jacob Weineri eksperiment liigiga kolmevärviline lehtertapp
(ronitaim), kellel on lihtne lahutada juur- ja võsukonkurentsi. Osad
taimed kasvasid üksinda potis, ilma igasuguste konkurentideta.
Teises seerias oli konkurente ühe kepi küljes mitu, kuid igal taimel
oli omaenda pott juurete kasvatamiseks. Kolmas seeria oli
vastupidine: kõigil oli ühine juurepott, kuid varred said igaüks oma
isikliku kepi najal kasvada.
Sümmeetrilise konkurentsi puhul (taimedel on selleks
juurkonkurents) jaguneb saadav kahju konkurentide vahel ligikaudselt võrdselt –
interaktsioon mõjub ühteviisi pärssivalt mõlemale osapoolele. Asümmeetrilise konkurentsi
puhul üks osapool osutub selgeks võitjaks, teine selgeks kaotajaks. Erinevalt
juurkonkurentsist on siin limiteeriv ressurss (PAR) varjestatav, s.t. kõrgem taim saab
madalamat efektiivselt varjutada, neelates enamuse pealelangevast kiirgusest.
KONSTANTSE SAAGI SEADUS JA ISEHÕRENEMISE SEADUS
Konstantse saagi seadust: olenemata külvitihedusest tuleb saak ikka samasugune, ainult siis
kui taimed jagavad ühist ressurssi, mitte ei kasva eraldi pottides või üksteisest kaugel. Saak
on ühe täiskasvanud taime keskmine mass (w) korrutatuna isendite arvuga pindalaühiku
kohta (N): w*N = const → w = const/N → w = cN-1 → log w = log c – log N.
Isehõrenemise seadus: lehepinna indeks (L) ei kasva lõputult koos taimede massi
suurenemisega vaid jääb mingist kohordi arengujärgust alates konstantseks. Isehõrenemise
ehk -3/2 astme seaduse matemaatiline väljendus: L = λN = const → λ = aD2 → w = bD2 → w
= b(L/a)2/2 N-3/2 → w = cN-3/2 → log w = log c - 1,5 log N, kus λ tähistab ühe täiskasvanud
taime keskmist lehepindala, D on taime poolt hõlvatud pindala diameeter, a ja b on
liigiomane konstant.
4. SÜNÖKOLOOGIA
POPULATSIOONIDEVAHELISTE INTERAKTSIOONIDE LIIGITUS
[o o] – neutralism ehk olulise vastasmõju puudumine. Koosluses võivad üksteise lähedal
elavad liigid, kelle vastasmõju on nii kaudne, et selle tähenduse võib nulliks lugeda.
[+ +] – mutualism ehk kahepoolselt kasulik vastasmõju.
Käitumuslik kohastumus – kaks liiki evolutsioonis omandanud käitumismustri, mis
on mõlemale kasulik. Näide: Aafrikas elavate aborigeensete hõimude ja suure
meenäitaja koostöö metsmesilaste pesade leidmisel ja seal leiduva toiduks
tarvitamisel.
Põllumajandus – sellega tegelevad maakeral sipelgad ja inimene, pakkudes oma
toiduobjektidele sööta või väetist ja kaitstes neid vaenulike liikide rünnaku eest.
Näide: aedniksipelgad kultiveerivad pesas seenekultuuri, keda nad pidevalt toidavad
taimelehtede saadud massiga. Sipelgate tegevus lehetäide „karjatamisel“.
Tolmeldamise ja seemneleviga seotud mutualism - enamik liike kasutab siiski
tolmlemiseks loomade abi – zoohooria (seedimatud seemned) või seemnete sipelglevi
ehk mürmekohooria. Loomadele pakkuvad õies leiduv suhkrurikas nektar või ka
pesapaika. Näide: õie nektarivaru sisaldav osa – nektaarium – on spetsiifilise
ehitusega ning ligipääsetav ainult kindla nokakujuga koolibridele.
Sümbioos ehk kooselu - mõistetakse selle all intiimset mutualismi – olukorda, kus
mutualistid elavad pikka aega koos „samas kehas“. Sümbioosis on nimetatakse suuremat
peremeheks ja väiksemat sümbiondiks.
Peremehe keha pinnal. Näide: peremeestaime juurte pinnal elavad ektomükoriisat
moodustavad seened. Mükoriisa ehk seenjuur on taimede ja seente sümbioosi, kus
seened saavad taimedelt fotosünteesi käigus toodetud suhkruid ning taimed omakorda
hädavajalikke mineraalseid toiteelemente.
Peremehe kehaõõnes. Näide: herbivoorsete või omnivoorsete (segatoiduliste)
loomade, sooltorus elavad seedimist abistavad bakterid. Loom ei suuda toota taimede
tselluloosist rakukesti lõhustavat ensüümi nimega tsellulaas.Tsellulaasi omavad
bakterid teevad selle töö loomade eest ära.
Peremehe kudedes. Näide: kõik liblikõielised taimed elavad sümbioosis
mügarbakteritega, moodustades liblikõieliste juurtesse kolooniaid – nooduleid. Taim
eraldab bakterite energeetiliste vajaduste katmiseks fotosüntaate, mikroobid
varustavad peremeestaime lämmastikuga. Autotroofid (fotobiont – tsüanobakter,
mükobiont – rohevetikas) moodustavad lihheniseerunud seente ehk samblike tihedalt
põimunud seeneniitidest kehandi, tallus ehk rakis. Vetikad varustavad seent
fotosüntaatidega, seen loob aga vetikale soodsa elukeskkonna.
Peremehe rakkude sees. Peremehe rakus elavat sümbionti nimetatakse
endosümbiondiks. Näide: taimed on
kompleksorganismid, kus olulise töö teevad
ära endosümbiondid – mitokonder ja
kloroplast.
Peremehe kehaõõnes ja rakkude sees.
Näide: sümbiontideks on zooksantellid –
mikroskoopilised ainuraksed vaguviburvetikad, kes leiavad varju korallide kudedes ja
kehaõõntes, varustades neid fotosünteesil toodetud suhkrute ning hapnikuga.
[- -] – konkurents ehk kahepoolselt kahjulik vastasmõju.
Võitluskonkurentsis segavad konkurendid üksteise tegevusele otseselt vahele, võideldes
toidu või pesapaiga pärast, või püüdes üksteist jahimaadelt välja tõrjuda. Näide: kiskjate
kaklus toidu pärast on tavaline, taimede valguskonkurents.
Võistluskonkurents toimib läbi ühise ressursi ammutamise. Näide: nahkhiireliiki saagi
leidmisel, taimede juurkonkurentsis.
Kaudne konkurents toimub populatsioonide vahel, kes ühe ja sama ressursi peale ei
konkureeri, aga vahemeheks on kolmas liik, kes on kohastunud mõlema kaudse konkurendi
ressursse tarbima.
Näiv konkurents toimub läbi ühise vaenlase mõnel muul troofilisel tasemel. Näide:
valgesaba-pampahirvel ja põdral oli ajuparasiit, kes nakatas mõlemaid liike, kuid oli surmav
ainult põdrale. Hirv oli parasiidi edasikandja, mitte „aus“ konkurent.
G. Gause konkurentsieksperiment:
Georgi Gause korraldas eksperimendi pärmidest toituvate amööbidega perekonnast
Paramecium. P. caudatum suri, P. aurelia võitis, P. bursaria aga suutis konkurendiga eluruumi
ja ressurssi jagada. Nii P. aurelia kui P. caudatum toitusid vees hõljuvatest pärmirakkudest, P.
bursaria toitus põhja sadenenud pärmirakke.
Gause reegel: „Kui kahe liigi nõudlused limiteeriva ressursi suhtes kattuvad, ei saa nad pikka
aega koos eksisteerida, vaid üks tõrjutakse konkurentsis välja.“ Hutchinson konkurentse
väljatõrjumise seadus: „Kaks liiki, kelle ökoloogilised nišid kattuvad, ei saa pikka aega
koos eksisteerida vaid üks tõrjutakse konkurentsis välja.“ Kaks taksonoomiliselt lähedast
liiki, kas jagavad sama eluruumi, ei saa oma nõudluste poolest olla väga sarnased. See
tähendab, et eksisteerib piir ehk limiteeriv sarnasus, kuivõrra kokkulangevad võivad
rahumeelselt koos eksisteerivate liikide elukombed ja toidueelistused olla. Tugevast
konkurentsisurvest tulenevat ressursinõudluste lahknemist ja sellekohaseid muutusi liigi
morfoloogias, anatoomias või füsioloogias nimetatakse tunnuse nihkeks.
[+ -] – tarbimine (näkitsemine, röövlus, parasitism) ehk ühe populatsiooni kahjustamine
teise populatsiooni poolt.
Näkitsemise puhul tarbitakse saakorganismi osi ilma saaki tapmata. Näide: kõrrelistel
paikneb meristeemlehe alaosas, mis tähendab, et herbivooride hammustamisel lehe kasv ei
lakka, vaid leht pikeneb edasi. Meristeem on taimedele omane kude, mis vastutab taime
kasvu eest.
Röövluse puhul surmatakse saakorganism tarbimise eesmärgil rünnaku käigus. Loomtoidulisi
loomi nimetatakse kiskjateks ja nende saagihankimiskäitumist kiskluseks.
Kiskjad jagunevad saagi püüdmisel:
Varitsejateks (kalad, maod ja konnad);
Aktiivseteks saagiotsijateks: jälitajaid (suudavad saagi jälitamise käigus ära
väsitada või saagist kiiremini liikuda, hundid või gepardid); sööstjaid (üllatavad
saaklooma kiire ja täpselt sihitud sööstuga, ämblikud, röövkalad või kullilised);
vastajaid, kes suudavad lõhna või mõne muu signaali abil peitunud saakorganismi
üles leida (elektrosensoorsed haid, rebased, rähnid); püüniste seadjaid (ämblikud,
herilased ja insektivoorsed taimed, inimene).
Saakloomade kaitsekohastumused kisklusele:
Pelgupaik;
Kiire liikumisvõime;
Keemiline kaitse (ebameeldiv või mürgine lõhn ja maits);
Regeneratsioonivõime;
Mehhaaniline kaitse (ogad või okkad, mis teevad tarbimise keeruliseks);
Värvuse ja kehamustriga seotud kaitsekohastumused:
Varjevärvus (selle abil imiteerib saakorganism oma elukeskkonna värve ja
mustreid);
Hoiatav värvus (silmapaistvalt ere ja kombineeritud keemilise kaitsega).
Mülleri mimikri ehk imiteerimine, kus mürgised või muidu mittesöödavad
liigid omandavad evolutsioonis ühise kiskjatele hästi ära tuntava
hoiatussignaali oma söödamatuse kohta. Batesi mimikri on kohastumus, kus
tegelikult ohutud mittemürgised liigid imiteerivad mürgiseid, et vähendada
kiskjarünnakuid ja seeläbi oma kohasust tõsta;
Ehmatav värvus ja kuju (kergesti märgatavatel liikidel on kohastumusi,
mille eesmärk on jätta kiskjale ekslik mulje palju suuremast ja ohtlikumast
loomast);
Segav värvus.
Lotka-volterra kiskja-saakloom võrrandsüsteem
, kus
dN/dt – kasvukiirus;
r – erikasvukiirus;
N – saaklooma populatsioonitihedus;
P – kiskja populatsioonitihedus;
a’ – efektiivsus, millega kiskja otsib ja tabab saaki;
f – koefitsient, mis näitab, kui efektiivselt kiskja tabatud saagi järglasteks
konverteerib;
q - liigiomane konkurentsikoefitsient.
Populatsioon on stabiilne ehk saaklooma ja kiskja populatsiooni kasvukiirus on null:
Saaklooma ja kiskja populatsioonid saavad korraga olla stabiilsed vaid väga piiratud
tingimustel. Igal ajahetkel on süsteem dünaamiline, mitte ideaalses nullkasvus – sõltuvalt
mõlema populatsiooni suurusest võivad mõlemad kiirused olla nii positiivsed kui negatiivsed.
Suur kiskjate hulk mõjutab saaklooma populatsiooni negatiivselt, aga suur saakloomade
tihedus mõjutab kiskja populatsiooni positiivselt.
Parasitismi ehk nugimise puhul on interaktsioonis kasu saav ning kahju kannatav organism
omavahel pikemat aega intiimselt seotud, parasiit saakorganismi ei tapa, vähemalt mitte
esimese rünnaku käigus.
Parasiitide liigitus:
Makroparasiitide hulka kuuluvad need, keda on mõistlik ja võimalik isendite kaupa
loendada.
Mikroparasiitide puhul hinnatakse parasiidi populatsiooni suurust nakatunud
peremeesorganismide arvu järgi.
Fakultatiivsed parasiitid ehk hemiparasiitid (poolparasiitid). Hemiparasiite on
roheliste lehtedega taimed, kes lisaks fotosünteesile tegelevad toidu- ja energiavarude
täiendamiseks ka parasitismiga. Hemiparasiidid nakatavad naabertaimede juuri,
kasvatades juuretipus asetseva spetsiaalse organi, haustori ehk imitoru, naabri juure
sisse ja imevad viimase koemahlu.
Obligatoorsed parasiitid ehk holoparasiitid (täisparasiitid). Holoparasiidid
iseseisvalt eluga hakkama ei saa, vaid vajavad elus püsimiseks ühte või mitut
peremeesorganismi. Kui järjestikuseid peremehi on mitu, nimetatakse varasemaid
vaheperemeesteks ja nugilise paljunemisfaasi-aegset peremeest pärisperemeheks.
Nende kehaehitus on tihti lihtne ja paljud aktiivse elu jaoks tarvilised organid
redutseerunud või kadunud.
Mükoheterotroofia on käpaliste sugukonnale iseloomulik pool- või täisparasitism.
Orhidee ja seene parasitism, kus orhidee omastab seene poolt omastatud orgaanilisi ja
mineraalseid toitaineid ilma fotosüntaate vastu pakkumata.
Nekrotroofsed parasiitid surmavad alati peremehe ja tegutsevad pärast seda edasi
kas ainult saprotroofina, algul saprotroofi ja hiljem näkitseja või kiskjana
(parasitoid), või siis ainult näkitseja või kiskjana.
Mikroparasiidi puhas kasvukiirus
Parasitoloogias on oluline teada, kas nakkushaigus levib, taandub või püsib nakatunute hulk
samal tasemel. Seda peegeldab parasiidi puhas kasvukiirus Rp, R0 näitab mitu järglast üks
keskmine populatsiooni liige elu jooksul. Parasiidi puhast kasvukiirust nimetatakse ka
nakatuskordajaks. Kui R0 = 1, siis nakatab üks haigestunud isend haiguse jooksul
keskmiselt ühe terve isendi ja nakatunute protsent püsib ajas muutumatuna. Kui R0 > 1 siis
nakkus levib, kui R0 < 1 siis nakkus taandub.
Parasiidi puhas kasvukiirus sõltub kolmest tegurist:
Mikroparasiidi ülekandekiirus (β)), st keskmine kiirus, millega parasiit ühelt isendilt
teisele levib. Ülekandekiirus sõltub esiteks parasiidi nakatamisvõimest ehk
infektsioossusest. Viimane näitab, kui hõlpsalt mikroparasiit nakatunud peremehelt
uuele peremehele üle kandub;
Aeg, mille jooksul peremeesorganism jääb nakatunuks (L), periood haiguse
summaarsest kestusest, mil nakatunu on parasiidi võimalik edasikandja;
Nakkusele vastuvõtlike peremeesorganismide populatsioonitihedus (S).
Parasiidi puhas kasvukiirus avaldub kolme ülalpool loetletud parameetri produktina: R0 =
β)LS. Vaktsiini olemasolul suudetakse mõjutada haigusele vastuvõtlike isendite
populatsioonitihedust (S). Karjaimmuunsuse tekkimisel parasiidi levik peatub. Sellist
nakkusele vastuvõtlike isendite populatsioonitihedust, mille puhul Rp = 1, nimetatakse
kriitiliseks tiheduseks (Skr): R0 = 1 → βLS = 1 → Skr = 1/βL. Minimaalse immuniseerimist
vajavate inimeste proportsiooni (pkr): pkr = 1 - Skr/S0.
[+ o] – kommensalism, ehk ühele poolele kasulik kuid teisele poolele kahjutu (neutraalne)
vastasmõju.
Paljudel juhtudel asustavad kommensaalid „märkamatult“ elukeskkonda, mis on vähemalt
osaliselt loodud mõne muu bioloogilise liigi poolt. Näited: samblikud ja samblad kasutavad
puutüvesid kasvusubstraadina, muutmata sellega puittaimede populatsioonitihedust, suur osa
loomade kehapinda asustavatest mikroobidest ei paku loomaliigile otseseid hüvesid, kuid ka
ei kahjusta neid.
Laguahel kui kommensalistlik süsteem
Laguahel on doonorkontrollitud süsteem ja sobib seepärast hästi kommensalismi
näiteks. Laguahelas osalevad liigid enamasti ei mõjuta otseselt toiduobjekti
populatsioonitihedust ja assimileerivad sellist surnud orgaanilist ainet, mis siseneb
laguahelasse neist mitte olenevail põhjusil. Surnud orgaanilisest ainest toituvaid
organisme nimetatakse
saprotroofideks
(saproobideks,
saprobiontideks,
saprofaagideks), st nad ei põhjusta kunagi otseselt toiduobjekti surma. Oraanilise
aine dissimilatsiooni viivad alati lõpule bakterid ja seened, keda seetõttu kutsutakse ka
tõelisteks lagundajateks. Laguahela esimesi liike, neid, kes toituvad morfoloogilise
struktuuri säilitanud surnud orgaanikast ehk detriidist, nimetatakse detritivoorideks.
[- o] – amensalism ehk ühele poolele kahjulik kuid teisele poolele neutraalne vastasmõju.
KOOSLUSTE STRUKTUUR
Taksonoomiline struktuur. Kõige olulisem aspekt on liigiline koosseis, annab aimu sellest,
millised liigid on kohastunud elama sarnastes keskkonnatingimustes, ja just sellises
keskkonnas, mis vaadeldavas ökosüsteemis leida on.
Ruumiline struktuur, kuidas ning milliste faktorite toimel varieerub koosluste liigiline
koosseis ja elurikkus ruumis. Olenevalt ökosüsteemi tüübist ja uuritava koosluse omapärast,
võib peamist huvi pakkuda kas üks ruumimõõde (vertikaalstruktuur), kaks ruumimõõdet
(horisontaalstruktuur), või ka kõik kolm ruumimõõdet (ruumiline strukrur).
Gradientanalüüsi käigus selgitatakse, millised koosluse struktuuri aspektid, ja mil moel,
korreleeruvad, ja on seletatavad, keskkonna erinevate mõõdetavate omadustega.
Ajaline struktuur. Sesoonsed muutused on seotud aastaaegade vaheldumisega. Lühiajaliste
muutuste taga on reeglina kliimatingimuste fluktuatsioonid või häiringud. Pikemaajalisi
avalduvaid suunatud muutusi koosluste struktuuris nimetatakse suktsessioonilisteks
muutusteks, protsessi ennast koosluse suktsessiooniks. Suktsessiooni võivad esile kutsuda nii
kooslusesisesed biootilised kui ka välised mõjurid – abiootiliste tingimuste muutumine väliste
tegurite mõjul või tugevad häiringud.
KOOSLUSTE DISKREETSUS VS PIDEVUS
Organitsistlikuks (klementsiaanlikuks) seetõttu, et Clements vaatles taimekooslusi kui
koevolutsioneerunud liikide kogumit, millel on nn superorganismi tunnused (tööjaotus,
vastastikune kasulikkus jne). Individualistlikuks (gliisoniaanlikuks) sellepärast, et Gleasoni
arvates kujutas kooslus endast lihtsalt liikide konglomeraati, kus elutu keskkonna isepärad ja
vastupanuvõime kohalikele vaenlastele ära nimekirja liikidest, kes hakkavad sama eluruumi
jagama. Organitsistlikust vaatest järeldub taimekoosluste diskreetne ehk hüppeline üleminek
ühest kooslustüübist teise nii ruumis kui ajas. Individualistliku vaate kohaselt muutuvad
kooslused keskkonnagradiendil pidevalt ehk kontinuaalselt – seoses abiootilise keskkonna
muutumisega kaovad järk-järgult ära mingid liigid ja asenduvad uutega. Organitsistliku
käsitluses on süntaksonoomiline süsteem, mille põhiüksuseks oli assotsiatsioon.
Individualistlikud arendasid välja taimekoosluste ordinatsiooni meetodid. Ordinatsioon
tähistab loomult keeruliste paljumõõtmeliste taimkatteandmete töötlemist viisil, mis lubab
kooslusi taksonoomilise sarnasuse/erinevuse järgi järjestada, korrastada ning võrrelda.
EESTI METSAKASVUKOHATÜÜBID E. LÕHMUSE JÄRGI
E. Lõhmuse metsakoosluste klassifikatsiooni põhiüksuseks
on
metsakasvukohatüüp.
Metsakasvukohatüübid
koonduvad
tüübirühmadeks, viimased omakorda
moodustavad kaks tüübirühmade klassi – arumetsad ja
soometsad. Arumetsade klassi kuuluvad parasniiskel või
mõõdukalt liigniiskel mineraalmullal mullal levivad metsad,
kus turbahorisont kas puudub, või selle tüsedus ei ületa
looduslikus olekus 30 cm. Soometsade klassi kuuluvad
liigniiskel mullal kasvavad metsad, kus turbahorisondi
tüsedus looduslikus olekus ületab 30 cm.
ARUMETSAD
Nõmmemetsad – kuivadel ja toitainetevaestel liivastel muldadel kasvavad metsad.
Eriti kuivades oludes levib sambliku kasvukohatüüp. Paremates niiskustingimustes
kohtab kanarbiku kasvukohatüüp.
Loometsad – paepealsetel neutraalse või aluselise reaktsiooniga kuivadel õhukestel
(<30 cm) kuid huumusrikkastel ja seega üpris toitaineterikastel muldadel kasvavad
metsad. Eriti õhukese mullakihiga (<12 cm) ja kergesti läbi kuivavatel muldadel levib
leesikaloo kasvukohatüüp. Veidi tüsedamatel muldadel levib kastikuloo
kasvukohatüüp. Lubikaloo kasvukohatüüp, mida kohtab paikades, kus on
vahelduv veerežiim, mis varieerub liigniiskest väga kuivani.
Palumetsad – tüübirühm, mis levib liivastel toitainete poolest vaestel või keskmistel
muldadel. Toitenetvaesem palumets kuulub pohla kasvukohatüüpi, toitaineterikkam
mustika kasvukohatüüpi.
Laanemetsad – keskmise viljakusega ja viljakal parasniiskel mullal kasvavad metsad.
Jänesekapsa kasvukohatüüpi kuuluvad metsad on ökoloogiliste tingimuste poolest
Eesti keskmised (keskmine mullaniiskus, keskmiselt viljakas muld). Veidi kuivemal
ja veidi lubjarikkamale lähtekivimile tekkinud mullal levib sinilille kasvukohatüüp.
Salumetsad – viljakad tüsedal ja huumusrikkal mullal kasvavad kuusikud või kuuse-
sega- või lehtmetsad, mille puurinne võib olla üsna liigirikas. Parasniiskel mullal
levivad salumetsad kuuluvad naadi kasvukohatüüpi. Ajuti liigniiskel mullal
kasvavad sõnajala kasvukohatüüpi salumetsad.
Soovikumetsad – perioodiliselt liigniisked turvastunud huumushorisondiga
rohttaimederikkad leht-(kask sanglepp) või segametsad. Veidi niiskematel ja veidi
toitaineterikkamatel muldadel levivad
osja kasvukohatüüpi
ja
tarna
kasvukohatüüpi kuuluvad soovikumetsad.
Rabastuvad metsad – neid iseloomustab liigniiske ja happeline muld. Sinika
kasvukohatüüpi liigituvates metsades on mulla lähtekivimiks mineroogiliselt
koostiselt vaesed liivad. Karusambla kasvukohatüüpi metsade mulla lähtekivimiks
on raske lõimisega savid ja liivsavid.
SOOMETSAD
Rohusoometsad – seal on turbahorisont moodustunud peamiselt rohttaimede ja
puittaimede jäänustest. Lodu kasvukohatüüpi kuuluvad metsad on peamiselt
kaasikud ja sanglepikud, mis kasvavad potentsiaalselt viljakatel hästi lagunenud
turbalasundiga muldadel. Madalsoo kasvukohatüüpi liigituvates metsades on
turbalasund enamasti keskmiselt lagunenud, puurindes domineerib sookask.
Samblasoometsad – neile metsadele on iseloomulik vähe lagunenud, peamiselt
turbasambla jäänustest koosneva turbakihi esinemine, vähemalt turbalasundi ülaosas.
Siirdesoo kasvukohatüübis
on mikroreljeef tugevasti künklik.
Raba
kasvukohatüübi metsades katab maad pidev turbasamblavaip, puhmarinne on hästi
välja arenenud.
Kõdusoometsad – need metsad levivad inimese poolt kuivendatud sooaladel, kus
aastatuhandete jooksul ladestunud turbalasund on põhjavee taseme alanemise tõttu
kõdunema hakanud. Mustika-kõdusoo kasvukohatüüp on nime saanud niiskema
mullaga palumetsa järgi, veidi viljakam jänesekapsakõdusoo kasvukohatüüp
omakorda niiskema mullaga laanemetsa järgi.
KOOSLUSTE SUKTSESSIOON
Suktsessiooniks nimetatakse ökosüsteemi keskkonnatingimuste ja sellest tulenevalt ka
ökosüsteemi eluskoosluse suunatud muutumist ajas. Muutused ökosüsteemis võivad olla
tingitud seda asustavate organismide elutegevusest – sel juhul on tegemist autogeense
suktsessiooniga. Näide: parasvöötme soode suktsessioon. Suktsessioonilisi muutusi võivad
esile kutsuda ka mingid välised faktorid, mis mõjutavad oluliselt keskkonnatingimusi ja
kutsuvad esile liikide vaheldumise eluskoosluses – sel juhul on tegemist allogeense
suktsessiooniga. Näide: Eesti soometsade kuivendamisel toimuvad muutused, mis lühemas
ajaskaalas päädivad kõdusoometsade tekkega.
Soode suktsessioon
Soo tekkimine algab enamasti madalaveelise ja seisva veega veekogu kinnikasvamisest.
Lagundajad tarvitavad vees lahustunud hapniku ära, laguprotsessid aeglustuvad ning surnud
taimeosad kuhjuvad veekogu põhja turbana. Esimene faas – madalsoo, siis kui turba
akumulatsioon jõuab kriitilise piirini, tekib veekogu pinnale õõtsik. Edasi eristuvad soos
selgelt oligotroofsed turbasamblased mättad ja eutroofsed madalsootaimedega asustatud
mätastevahelised lohud – siirdesoo faas. Turbahorisondi edasisel tüsenemisel oligotroofsed
turbasamblaga kaetud mättad liituvad ja kujuneb välja oligotroofne ja vihmaveetoiteline ehk
ombrotroofne – kõrgsoo ehk raba. Rabas jääb igal aastal lagundamata umbes 10% taimsest
biomassist ja turbakiht tüseneb selle arvelt ca 1 mm võrra aastas.
Kui suktsessioon algab seni taimestumata maapinnal või seni organismidega asustamata
veekogus, nimetatakse seda primaarseks suktsessiooniks. Näide: hangunud laavaväljad,
mere alt maakerke tõttu vabanenud maismaa, ammendatud karjäärid, või liustike taandumisel
jää alt vabanenud maapind. Olemasolevate ökosüsteemide arengut pärast tugevaid häiringuid
nimetatakse sekundaarseks suktsessiooniks. Näide: lageraielangid või metsatulekahjude
poolt laastatud alad.
Kliimakskooslus
on suktsessiooni kliimaomane
lõppstaadium. Monokliimaksi teooria, milliseid teid pidi
sarnase kliimaga ja ühise liigifondiga piirkonnas
suktsessioon ka ei kulgeks, lõpptulemus on alati sama.
Oligokliimaksi teooria käsitluse põhjal suktsessioon võib
viia mitme alternatiivse stabiilse lõppstaadiumi suunas
sõltuvalt mulla lähtekivimist ja mulla niiskusrežiimist.
BIOLOOGILINE MITMEKESISUS
Elurikkuse tulipunktid on piirkondi maailmas, kus
elurikkus on võrreldes ülejäänud samasse kliimavööndisse
kuuluvate aladega anomaalselt suur. Valdavalt leiab selliseid
liigirikkaid ökosüsteeme niiskest troopikast, aga ka neis
parasvöötme piirkondades, mida iseloomustab vahemereline
kliima. Sellist kliimat kohtab 30. ja 45. laiuskraadi vahel,
peamiselt kontinentide lääneservades.
Suuremal skaalal on rekordilised taimede elurikkuse numbrid loetud Kesk-Ameerika ja
ekvatoriaalse Lõuna-Ameerika troopilistes vihmametsades. Teataval skaalal (paar
ruutdetsimeetrit) on taimekoosluste liigirikkuse maailmarekordid registreeritud Eestis,
Läänemaal Virtsu lähistel paikneval Laelatu puisniidul.
Mitmekesisuse globaalsed gradiendid
Laiuse gradiendil. Ökosüsteem asustavate liikide arv on reeglina suurim ekvaatori
läheduses ja väheneb pooluste suunas.
Absoluutse kõrguse ja sügavuse gradiendil. Absoluutne kõrgus on negatiivselt
seotud aasta keskmise temperatuuriga. Seega elurikkus absoluutse kõrgusega reeglina
langeb. Veekogudes elurikkus sügavusega üldjoontes langeb mite päris monotoonselt.
Primaarse produktiivsuse gradiendil. Mida produktiivsem maismaa- kui
veeökosüsteem, seda rohkem liike seal elab. Ökosüsteemide primaarset produktiivsust
saab üpris hästi ennustada läbi potentsiaalse evpotranspiratsiooni (PET). PET on
kliimaomane mõõt, mis näitab kui palju vett potentsiaalselt ökosüsteemis mingi
ajaühiku jooksul maapinnaühiku kohta aurub (evaporatsioon) ja läbi taimede mullast
atmosfääri liigub (transpiratsioon).
Mitmekesisuse lokaalsed gradiendid
Rannikulähedastes ökosüsteemides kipub liigirikkus üldjuhul vähenema poolsaare
otsa või lahesopi suunas, võrreldes sisemaa või avaveekoguga – poolsaare efekt
(maismaal) või lahe efekt (veekogus). Põhjus: meres olevasse suvalisse punkti võivad
vaadeldava keskkonna potentsiaalsed asukad e liigifondi liikmed migreeruda igast
küljest. Lahte saavad nad liikuda ainult ühest suunast, sest laht on piiritletud
maismaaga.
„Küürselg-kõver“ – maksimaalse liigirikkuse leiame mingi keskmise või väikese
produktiivsuse korral, väga vaesed ja väga produktiivsed kooslused on liigivaesed.
Liikide arvu ja pindala seos
logS = logc + zlog A → S = cAz (S – liikide arv proovipinnal, A – proovipinna pindala, c ja z
– võrrandi koefitsiendid ehk parameetrid). Juhusliku või regulaarse ruumile jaotusega
populatsioonide segust koosnevas koosluses on z maksimaalne. Kui kooslus koosneb suurtest
monokultuure moodustavatest populatsioonidest, on z minimaalne (maismaa Z=0,09; saarel
Z= 0,30).
Elurikkuse varieerumine ajas
Elurikkuse lühiajalised muutused ehk aasta-aastased muutused on seotud
temperatuurirežiimi ja sademete hulga varieerumisega aastast aastasse.
Elurikkuse suktsessioonilased muutused ehk mõnekümne kuni mõne tuhande
aastani muutused on väga varieeruvad ning neid iseloomustab alati liikide
vaheldumine ajas ja sellega kaasneb enamasti liigirikkuse tõus või langus kindlates
suktsessioonijärkudes.
Elurikkuse muutumine evolutsioonilises ajas sõltub liigitekke ja liikide
väljasuremise dünaamikast vaadeldavas organismirühmas ja pikaajalistest
kliimamuutustest, mis määravad ära ühe või teise bioomi olemasolu paiknemise ning
pindala maakeral. Laias laastus on elurikkus evolutsioonilises ajas suurenenud.
Massväljasuremisteks ehk massekstinktsioonideks on episoode, kus suurem osa
maakera liikidest lühikese aja jooksul välja sureb. Tänapäeval on käimas
antropotseeni massekstinktsioon, mis on põhjustatud inimese tegevusest maakeral.
ELURIKKUSE HINDAMINE JA MÕÕTMINE
Diversiteedi komponendid: liigirikkus, mis näitab liikide arvu pindalaühikul; ühtlus, mis
mõõdab, kui ühtlaselt on ressursid erinevate taksonite vahel jagunenud.
Diversiteediindeksid, mis hindavad mõlemat diversiteedi komponenti – liigirikkust ning
ühtlust.
Simpsoni indeks (ʎ) on „antidiversiteediindeks“, st see hindab küll üheaegselt nii
liikide arvu kui ühtlust, kuid teeb seda tagurpidi – mida suurem on indeksi väärtus,
seda väiksem on süsteemi mitmekesisus. Simpsoni indeks mõõdab tõenäosust, et kaks
kooslusest juhuslikult valitud isendit on ühest ja samast liigist. Valem: λ= Σ pi2;
Shannoni indeks (H’) näitab korrapäratust e entroopiat. Liigi osakaal on korrutatud
naturaallogaritmiga liigi osakaalust H´ = - Σ pilnpi;
Hilli perekonna indeksid (Na).
Ühtluse indeksiteks (E) soovitatakse erineva
ühtlusetundlikkusega Hilli indeksite suhteid. E2,0: E2,0 = N2/N0.
5. EVOLUTSIOONILINE ÖKOLOOGIA
Püütakse ökoloogiliste terminitega ökoloogilises taustsüüsteemis uurida organismide
kohastumusi, nende tekkimine, püüsimine ja kadumine ajas.
ELUKÄIGUTUNNUSED
Elukäigutunnused ehk kohasusega seotud tunnused:
eluiga (väikeealine, pikaealine), paljunemispingutuse arv
elujooksul,
keha suurus,
vanus suguküpsuse
saavutamisel.
TUNNUSE
EVOLUTSIOON
KUI
OPTIMUMIÜLESANNE
Optimaalne tähendab, et annab oraganismile kõige
suurema kohasuse.
Optimaalne kurna suurus on määratud sellega, millise
kurna suuruse puhul on paljunemise hinna ja järglaste
summarse kohasuse vahe kõige surem (pikkem). See on
keskmisest mõnevõrra väiksem.
LÕIVSUHETE JA ALLOMEETRILISTE
SEOSTE KUJUNEMINE
Lõivsuhe märgib negatiivseid seoseid organismi,
populatsiooni, taksoni erinevate võimekuste vahel.
Allomeetriline seos märgib positiivseid seoseid organismi, populatsiooni, taksoni erinevate
võimekuste või omaduste vahel.
Ressursside suutlikkus ja allokatsioon on määratud geenide abil. Sõltuvalt sellest, kus on
varieeruvus, kas lokuses A või lokuses B, kujuneb välja kas allomeetriline seos või lõivsuhe.
KÄITUMISÖKOLOOGIA
Käituja: kanakull – absoluutselt kõik ressursid, tuvi –
suhtub rahumeelselt, ilma konfliktideta.
Strateegia: null summa mäng – ressursse on lõplik hulk,
võidab see, kes suurema osa haarab; võit-võit strateegia –
pühendatakse uute ressursside loomisele või ressursside
säästlik kasutamine.
Altruism – enesele kahjulik käitumine mõne teise isendi seisukorra parandamiseks.
Vorst – vorsti – vastu altruism on sotsiaalne käitumismuster, kus investeering isetuse
käitumise tasub varem või hiljem ära, niikaua kuni sellega tegelevad kõik sotsiumi
liikmed;
Altruism lähisugulaste suhtes.
Hamiltoni reegel: altruism on evolutsiooniliselt mõistlik (võimalik) siis kui: enesele
tekitatud kahju < teis(t)ele tekitatud kasu * sugulusaste (r).
Sugulusaste (r): kaksikõve – r = 1 („õve“ tähendab õde-venda), õve/ema/isa – ½;
tädi/onu/vanaema/vanaisa – r = ¼, nõbu/vanatädi/vanaonu – r = 1/8.
Kaasav – kohuse printsiib: kui sa abistad mitte ainult iseennast, vaid kedagi, kes jagab
sinuga olulisel määral geene.
SUGULINE VS KLONAALNE PALJUNEMINE
Suguline paljunemine annab pidevalt uut ja kasulikku materjal lodusliku valikule.
Tulemuseks on geneetiliselt unikaalsed järglased (raiskav süsteem – efektiivne).
Fisheri reegel – 0,5:0,5 reegel – isaseid ja emaseid sünnib populatsioonides üldjuhul
võrdselt.
* Ühiselulised putukad – haplodiploidia – soo määrab haploidsus (isased – haploidsed,
emased – diploidsed).
SOO MUUTUMINE ELUEA JOOKSUL
Lahksugulisus – kindel soo.
Hermafrodism – mõlemasoolisus – organismis nii
ema, kui ka isa suguorganid.
Seriaalne hermafrodism – organism võimeline
arendama välja mõlemad suguorganid. Nad ei ole
korraga, vaid sugu võib vahetada elu jooksul.
SUGULISE VALIKU TÜÜBID
Otsene võitlus isaste vahel, mis võib toimuda, või edavõrdse võimaluse korral ka ära jääda
(heidutamine). Näide: loomaailmas on olemas rituaalid, kus
toimub isaste vahel kaklus relvlade (sarved, kihvad) abil;
Võistlus isaste vahel, kui isased otseselt üksteise kallal ei
käida, vaid püütakse emadele muljet avaldada mingisuguse
tegevusega. Näide: isaste lehtlalinnu meisterdatud lehtlad,
tundarapüüde mäng;
Händikäp-käitumine
ja
händikäp-tunnused,
enesele
kahjulik käitumine või enesele kahjulikud tunnused, mille
demonstreerimine tõestab võimekust ning häid geene. Võimekam ja parema tervisega isend
saab omale lubada käitumist või tunnuseid, mida teised ei ole. Händikäp ehk ede näitab
kellegi edumaad või üleolekut sooritusel. Näide: pabulinnu saba, lõvi lakk;
Keemiline valik on sugupooled eritavad teatus spetsiaalsed kemikaalid ehk feromoonid,
mille mõtte on signaliseerida oma kohalolust ja geneetilistest kvaliteedist. Näide: feromoonid
toodavad putukad, inimene;
Valik suguraku tasemel, sel juhul paaritub emane organism valimatult erinevate isastega ja
tehakse valik nende suguraku tasemel, mis on varutud. Näide: puuviljakärbes.
6. SÜSTEEMÖKOLOOGIA
ÖKOSÜSTEEMIDE PRODUKTIIVSUS
Ökosüsteemide produktiivsust mõõdetakse läbi energia või aine koguse, mis foto- või
kemosünteesi käigus salvestub elusainesse pindalaühiku kohta ajaühikus.
GPP – koguproduktsioon ehk summaarne primaarproduktsioon.
NPP – puhas primaarproduktsioon. NPP = GPP – R, kus R on taimne hingamine ehk
taimede enda elutegevuseks kuluv osa. Rusikareegli järgi kulub taimseks hingamiseks
ligikaudu pool kogu assimileeritud PAR-ist, NPP ≈ GPP/2.
Maismaa NPP = 115*109 t. Ookeanides NPP = 55*109 t,
maismaa NPP on ookeanide omast kaks korda suurem.
Ookeanides on suurim mandrite äärealadel (>90) ja väikseim
suurte ookeanide keskosas (< 35). 99,8% kogu biosfääri
kuivbiomassist asub maismaal, ookeanis vaid 0,2%, kuna
ookeanis esmane toodang on fütoplankton, kelle eluiga on
väga lühike. Troopilistesse vihmametsadesse, tänu
puittaimede pikale elueale, on 42% maismaa biomassist.
Parasvöötme metsade kogupindala on suurem kui troopilistel
vihmametsadel, kuid leiab sealt 34% maismaa biomassist.
Parasvöötme rohumaade ja savannide NPP opanus maismaa
globaalsesse biomassi on tühine (4%).
ENERGIA JA AINE LIIKUMINE LÄBI TROOFILISTE TASEMETE
Ökoloogiline püramiid ehk troofiline püramiid (Eltoni püramiid) on püramiid, mille
tasemed tähistavad sarnase toiduallikaga organismides ajaühikus toodetud orgaanilise aine
hulka või siis nendel troofilistel tasemetel igal ajahetkel immobiliseeritud orgaanilise aine
kogust ehk biomassi. Püramiidi kõige madalama astme vastab autotroofide ehk
primaarprodutsentide toodangule, teine aste esimest järku tarbijate toodangule, kolmas aste
teist järku tarbijate toodangule, kõige kõrgemal troofilisel tasemel paiknevaid tippkarnivoorid
(tippkiskjad). Parasiitide panust troofilise taseme toodangusse arvestatakse
peremeesorganismi toodangu hulka.
Toiduahel on aine ja energia liikumist läbi troofiliste
tasemete (autotroofid – herbivoorid – karnivoorid –
tippkarnivoorid). Toiduvõrgustik on toitumissuhete võrk,
kogum biotsönoosis või bioomis põimuvaid toiduahelaid.
Pn ja Pn-1 koguproduktsioon ja sellele eelneval troofilsel
tasemel. Alumisel tasemel salvestatud energiast ja ainest
jääb suur osa enne organismide surma tarbimata ja suundub otse laguahelasse. Ülejäänu on
sisendiks n-da troofilise taseme jaoks In. Osa sisendist väljutatakse alati fekaalse kaona Fn,
mis suundub otse laguahelasse. Organismi poolt omastatud osa moodustab assimileeritud
energia An. Kogu assimileeritud energiat ei saa organismid paraku kasutada produktsiooniks,
sest osa kulub elutegevuseks ehk rakuhingamiseks Rn või kasutatakse ära
bioproduktsiooniks Pn.
Tarbimisefektiivsus CE = In/Pn-1 – näitab kui efektiivselt suudetakse eelmisel
troofilisel tasemel salvestatud energiat tarbida;
Assimilatsiooniefektiivsus AE = An/In – näitab, millise efektiivsusega suudetakse
tarbitud toitu assimileerida;
Produktsiooniefektiivsus PE = Pn/An –
näitab, milline osa assimileeritud energiast
suudetakse produktsiooniks pöörata.
Troofiliste tasemete vahelise energia
ülekande efektiivsus TE = CE x AE x PE
= Pn/Pn-1. Globaalse üldkeskmise TE
hinnang 10%, siseneb biosfääris otse
laguahelasse
90%
immobiliseeritud
energiast.
Assimilatsiooniefektiivsus (A/I) sõltub peamiselt
toidu kvaliteedist, 80% lihatoidulistel loomadel ja
20% detriidist toitujatel. Produktsiooniefektiivsus (P/A) seletub peamiselt loomade
taksonoomilise kuuluvusega. Selgrootutel on hingamiskaod võrreldes selgroogsetega oluliselt
väiksemad ja PE seetõttu kordades suurem. Kõigusoojased selgroogsed (kalad, kahepaiksed,
roomajad) vajavad kasvamiseks ja paljunemiseks oluliselt vähem toitu kui püsisoojased
selgroogsed (linnud või imetajad), seega kasutavad püsisoojased 2% assimileeritud energiast,
kõigusoojaste 10%.
GLOBAALSED AINERINGED
Fond on koht ja seda iseloomustab aine kogus, mis seal keskmiselt igal ajahetkel leida on.
Voog kirjeldab elemendi või ainet ümberpaiknemise keskmist kiirust erinevate fondide vahel.
Süsinikuringe (1012 kg C)
Maakeral paikneb ringes olev süsinik peamiselt neljas
suures fondis: ookeani fond (38 000 ühikut), mulla
fond (1 500 ühikut), atmosfääri fond (720 ühikut),
maismaataimede maapealse osa fond (560 ühikut).
Süsinikuringe vood on mineraalse süsiniku liikumine
taimedesse fotosünteesil (120 ühikut maismaal ja 107
ühikut ookeanis) ning süsiniku vabanemine
süsihappegaasina
atmosfääri
ja
veekogudesse
aeroobse rakuhingamise käigus. Jõed toovad
ookeani aastas 0,2 ühikut süsinikku. Atmosfääri lisab
olulisel määral süsinikku ka metsade raie ja
metsamaa kasutusele võtmine põllumaana (2 ühikut aastas). Atmosfääri fondist viib
süsinikku välja lagundamata jäänud orgaaniliste setete ladestumine ookeani põhja (0,1
ühikut aastas) ja turba ladestumine soodes. Maailmamere süsinikuvaru ja atmosfääri fond
tänapäeval suureneb (fossiilse süsiniku kaevandamise ja oksüdeerimise arvelt, 5 ühikut).
Lämmastikuringe (109 kg N)
Bioloogilise lämmastikuringe oluline fond on
atmosfäär (78,1% koostisest). Bioloogiline N2
fikseerimine toob bioloogilisse ringesse 170
ühikut lämmastikku (18% toimub veekogudes).
Läbi
taimedele
omastatavate
lämmastikuühendite tekke
atmosfääris
toimuvate elektrilahenduste toel satub alla 10%
lämmastikku bioloogilisse ringesse. Primaarse
produktsiooni käigus assimileeritav lämmastik
pärineb ökosüsteemide sisesest ringest (1 200
ühikut maismaal ja 6 000 ühikut ookeanis). Kui laguahelas vabanenud mineraalset
lämmastikku kiiresti eiassimileerita, leostub see muldadest kergesti välja ja liigub põhjavette
või vooluvetega veekogudesse (10 ühikut lämmastikku ookeanisetetesse). Denitrifikatsiooni
teel lahkub atmosfääri tänapäeval rohkem lämmastikku (240 ühikut) kui seda bioloogilise ja
keemilise fikseerimise käigus taimedele kättesaadavaks tehakse (190 ühikut). Inimtegevus
(keemiatööstus, toiduainetetööstus, põllumajandus: karjakasvatus → ammoniaak) paiskab
lämmastikku atmosfääri (100 ühikut aastas). Inimese poolt õhulämmastiukst
lämmastikväetiste tootmine (42 ühikut). Ökosüsteemides ringleva pidevalt väheneb
lämmastiku kogus. Põhjuseks on siin inimtegevus –kultiveeritakse vähem lämmastikku
fikseerivate mikroobidega sümbioosis elavaid taimi.
Ammonifikatsioon (ammonifitseerijad bakterid) on aminorühma (-NH2) sisaldavate
orgaaniliste ainete bakteriaalne lagundamine. Ammonifikatsiooni lõpp-produkt on
ammoniaak (NH3). Nitrifikatsioon
(nitrifitseerijad bakterid) on ammooniumiooni
kaheastmeline oksüdeerimine, algul nitritiooniks (NH4+ → NO2-), pärast nitraatiooniks (NO2-
→ NO3-). Denitrifikatsioon (denitrifitseerijad bakterid) on mineraalne hingamine, milles
orgaanilise aine vesinikult võetavate elektronide lõppaktseptor on nitraat (NO3- ), mis
redutseeritakse kas dilämmastikoksiidiks (N2O) või molekulaarseks lämmastikuks (N2).
Fosforiringe (109 kg P)
Fosfororiringe omapäraks on atmosfääri fondi
puudumine, sest looduslikke fosforit sisaldavaid
gaase atmosfääris pole.
Fosforiringes
domineerivad neli peamist protsessi:
1)
tektooniliste protsesside käigus paljastuvad fosforit
sisaldavad kivimid, mille pinnakihtide murenemisel
saavad fosforit sisaldavad mineraalid
kättesaadavaks; 2) füüsilise erosiooni ja porsumise
tulemusena lahustuvad vees fosforirikkad mineraalid, mis satuvad mullavette ning
veekogudesse; 3) voolu- ja põhjavetega kandub fosfor järvedesse ja edasi jõgesid mööda
ookeani; 4) orgaanilistesse ühenditesse ja oksiididesse või karbonaatsetesse mineraalidesse
seotud fosfori settimine ja mattumine veekogude põhjasetetesse.
Veeringe (1012 kg H2O)
Ookeanist, kus paikneb 97% maakera veest, aurab igal
aastal atmosfääri 0,03%. Vähem kui 10% ookeanist
aurustunud veest kantakse maismaa kohale, kus see
lisandub transpiratsiooni ning evaporatsiooni käigus
ökosüsteemidest atmosfääri paisatud veeaurule ja langeb
sademetena maapinnale. Vooluveed viivad maismaalt
järvede ja jõgede kaudu pidevalt tagasi ookeani sama palju
vett kui seda ookeanist aurustub. Antartika, Gröönimaa ja
mäestike jääkilpidesse on immobiliseeritud ca 2%
maakera veest, põhjavees paikneb suhteliselt liikumatuna veel 0,6%.
Sellest failist leiad vastused ökoloogia kordamisküsimustele. Näiteks…
Põhimõisted - ökoloogia, biotsönoos, kooslus, ökosüsteem, biogeotsönoos…
Autökoloogia - PAR-i omadused, erineva lainepikkusega PAR-i saabumine päikeselt, klorofülli ja lisapigmentide neeldumisspektrid…
Demökoloogia - populatsioonitihedus, populatsiooni kasvukiirus, Malthuse võrrand, keskkonnatakistus…
Sünökoloogia - neutralism, mutualismi tüübid (näited), mutualsitlike sümbiontide
paiknemine peremehe keha suhtes (näited)…
Evolutsiooniline ökoloogia - elukäigutunnused (näited), tunnuse evolutsioon kui optimumiülesanne, lõivsuhete ja allomeetriliste seoste kujunemine…
Süsteemökoloogia - GPP ja NPP, produktiivsuse mustrid maismaal ja ookeanis, erinevate bioomide produktiivsus…
Sarnased õppematerjalid
![Ökoloogia kursuse konspekt]()
78
pdf
Ökoloogia kursuse konspekt.
Saab selgelt eristada, kust organism algab ja
kust lõpeb.
2.2 Modulaarne organism – keeruline. Organism koosneb moodulitest, mis on
suuremal / vähemal määral iseseisvad.
Nt: maikelluke
Ramet moodustab geneti.
Nt: harilik kuusk. Kuuse oks – 1 tüvest lähtuv oodul. Kui oks end ise ei suuda ülevad pidada
(on varjus/kahjustunud) siis lastakse oksal ära kuivada.
3. POPULATSIOONI TASE – Demograafiline ökoloogia
Isendid samast bioloogilisest liigist moodusavad populatsiooni. Üksi isendid hakkama ei saa,
ja kui saavadki – surevad varsti vlja.
Populatsioon...
1) ... vabalt ristuvate isendite kogum
DEEM – ehk pammiktiline populatsioon
2) ...mingi liigi isendid mingil ajal mingis kohas.
Operatsiooniline populatsioon
Populatsioon võib seega olla ka klassiruum, kus toimub info vahetus, puudub aga vaba
ristumine.
![ÖKOLOOGIA eksami küsimuste vastused]()
13
doc
ÖKOLOOGIA eksami küsimuste vastused
ÖKOLOOGIA (LOOM .01.105) KORDAMISKÜSIMUSED, kevad 2011. a.
1. Ökoloogia aine, alajaotused;
Teadus, mis käsitleb organismide ja keskkonna suhet. Kõikide sidemed kõikidega.
Jaguneb:
a) Ökofüsioloogia e molekulaarne ökoloogia
b) Autökoloogia (isendi/organismi tasandil)
c) Demökoloogia (populatsiooni tasandil)
d) Sünökoloogia (eluskoosluse, populatsioonide tasandil)
e) Süsteemökoloogia (ökosüsteemi tasandil, elus kooslus + eluta keskkond)
f) Biosfäroloogia e biosfääri ökoloogia (globaalne ökosüsteem)
2. Ökoloogia põhimõisted isend (genet, kloon, ramet), populatsioon, kooslus, ökosüsteem,
bioom;
Isend- kindla genotüübiga organism
![Ökoloogia konspekt]()
13
doc
Ökoloogia konspekt
1. Aine, alajaotused (allpool) , areng.
Ökoloogia - teadus, mis uurib elusa ja eluta looduse omavahelist suhet, ei keskendu ühele
objektile, vaatleb tervikut.
E. Haeckel 1869 ökoloogia on teadus organismide ja kk suhetest.
E. Odum teadus looduse struktuurist ja funktsoonist.
2. Ökoloogia põhimõisted.
Ökoloogia valdkonnad:
1) Organelli tase
2) Raku tase (ainurakse puhul isend)
3) Koe tase
4) Organi tase
5) Isendi tase autökoloogia, uurib abiootilisi kk faktoreid.
6) Populatsiooni tase demökoloogia e. populatsiooni ökoloogia.
7) Koosluse tase kooslusökoloogia e. sünökoloogia, uurib mitmeliigilisi pop. süsteeme.
![Nimetu]()
42
docx
Nimetu
72. Kuidas avaldub produktsiooniefektiivsus?
A. PE = In/Pa-1
B. PE = An/In
C. PE = Pn/An-1
D. PE = Pn/An
73. Kas kiskja-saaklooma süsteemis:
A. Kiskja populatsioonitiheduse madalseis vastab saaklooma populatsiooni suurimale tihedusele
B. Kiskja ja saaklooma populatsioonitihedus võnguvad samas rütmis
veerandperioodilise faasnihkega
C. Saaklooma populatsioonitihedus on kiskja omast sõltumatu kuid fluktueerib samas rütmis
74. Termini ökoloogia võttis kasutusele:
A. K. Zobel
B. V. Masing
C. Haeckel
D. Hutchinson
E. V. Gause
75. Seen võib olla:
A. Loom
B. Imetaja
C. Nematood
D. Parasiit
E. Primaarne produtsent
76. Produktsiooni efektiivsus on suurem:
A. Kõigusoojastel selgroogsetel?
B. Püsisoojastel selgroogsetel?
C. Nendel selgroogsetel, kelle energiakadu hingamisele on null?
77. Millise elemendi ringe puhul on atmosfäär väheoluliseks fondiks?
A. N
B. P
C. C
D. O
78
![Ökoloogia kordamisküsimused]()
11
doc
Ökoloogia kordamisküsimused
1. Aine, alajaotused (allpool) , areng.
Ökoloogia - teadus, mis uurib elusa ja eluta looduse omavahelist suhet, ei keskejdu ühele objektile,
vaatleb tervikut.
E. Haeckel 1869 - ökoloogia on teadus organismide ja kk suhetest.
E. Odum - teadus looduse struktuurist ja funktsoonist.
2. Ökoloogia pôhimôisted.
Ökoloogia valdkonnad:
1) Organelli tase- uurib olulisi eluavaldusi madalamal str tasemel
2) Raku tase (ainurakse puhul isend)
3) Koe tase
4) Organi tase- autökoloogia, org. Ja keskk. Suhete uurimine isendi tasandil
5) Isendi tase - autökoloogia, uurib abiootilisi kk faktoreid.
6) Populatsiooni tase - demökoloogia e. populatsiooni ökoloogia.
![ÖKOLOOGIA kordamisküsimuste vastused 2012]()
18
doc
ÖKOLOOGIA kordamisküsimuste vastused 2012
1. Ökoloogia aine, alajaotused;
Teadus organismide ja nende keskkonna vahelistest seostest (biootiline, abiootiline KK)
1) Molekulaarne ökoloogia (molekul) meetodile viitav.
2) Ökofüsioloogia (molekul, organ, isend) uurib füsioloogiliste protsesside kohanemist vastavalt keskkonnale.
3) Antökoloogia (isend) isendi suhted keskkonnaga.
4) Populatsiooniökoloogia, demökoloogia(demograafiline) (populatsioon)
5) Sünökoloogia, kooslusökoloogia (kooslus)
6) Süsteemökoloogia (ökosüsteem)
7) Geograafiline ökoloogia (bioom)
8) Biosfäroloogia (biosfäär)
2
![ÖKOLOOGIA kordamisküsimused 2013]()
11
doc
ÖKOLOOGIA kordamisküsimused 2013
ÖKOLOOGIA (LOOM .01.105) KORDAMISKÜSIMUSED, kevad 2013. a.
1. Ökoloogia aine, alajaotused;
Ökoloogia on teadus organismide ja keskkonna vahelistest suhetest.
Ökoloogia alajaotused on :
· Ökofüsioloogia (organell, rakk, organ)
· Autökoloogia (isend) organism ja keskkonna suhe isendi tasemel
· Demökoloogia (populatsioon)
· Sünökoloogia (kooslus)
· Süsteemökoloogia (ökosüsteem, biosfäär)
2. Ökoloogia põhimõisted isend (genet, kloon, ramet), populatsioon, kooslus, ökosüsteem, bioom;
· isend organism, mis ei moodusta iseseisvaid mooduleid
o kloon ehk genet geneetiliselt identne moodulite kogum
o ramet taime puhul võsu, risoomiga ühendatud, iseseisva juuresüsteemiga moodul
· populatsioon ühise genofondiga isendite kogum
· kooslus koos eksisteerivad populatsioonid
· ökosüsteem elukooslus ja selle abiootiline kes
![Ökoloogia eksami kordamisküsimused]()
15
docx
Ökoloogia eksami kordamisküsimused
Ökoloogia on teaduslik õpetus organismide ja nende keskkonna
vahelisest seostest ja mõjudest. Keskkonna alla kuulub nii biootiline kui ka abiootiline keskkond.
Ökoloogiat võib defineeida ka kui organismide "kodu elu". Ökoloogia alajaotused: * molekulaarne
ökoloogia (molekuli, organi ja isenfi tasandil) ; (ökofüsioloogia- uurib organismide
kohanemisreaktsioone) * autökoloogia (isendi tasandil) * pop.ökoloogia e demökoloogia *
kooslusökoloogia e sünökoloogia *geograafiline ökoloogia * biosfääriline ökoloogia
2. Ökoloogia põhimõisted isend (genet, kloon, ramet), populatsioon, kooslus, ökosüsteem,
bioom (konspekt);
Isend: unitaarne organism. Selline organism, kes ei moodusta mooduleid, mis oleksid kas suhteliselt
või täiesti iseseisvad.
Populatsioon: ühise genofondiga isendite kogum.
Kooslus: koos eksisteerivad populatsioonid
Ökosüsteem: hõlmab endas elukooslust ja selle abiootilist keskkonda
Bioom: saransed ökosüsteemid üle maailma
3
Meedia
Kommentaarid (0)












































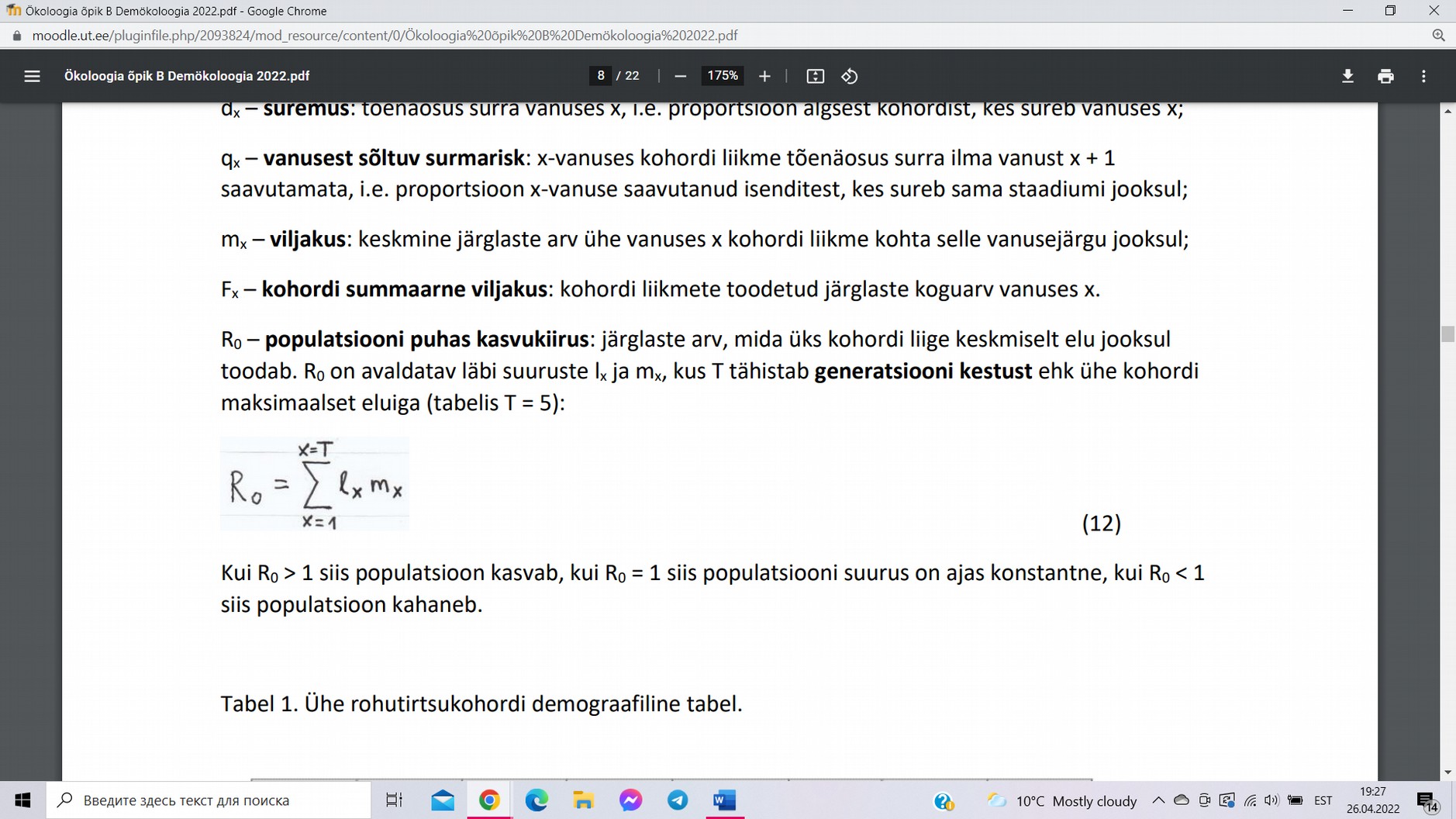
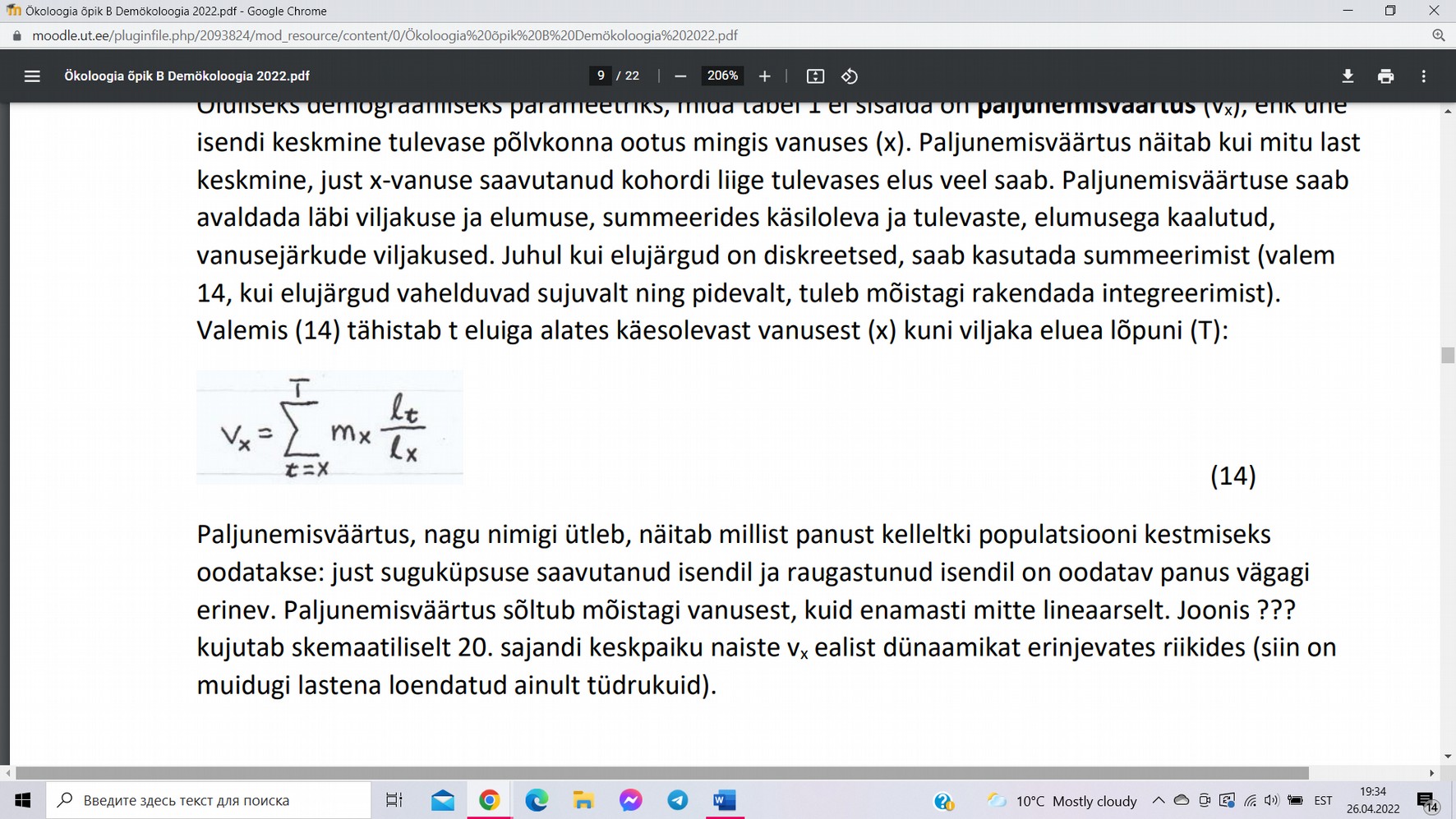
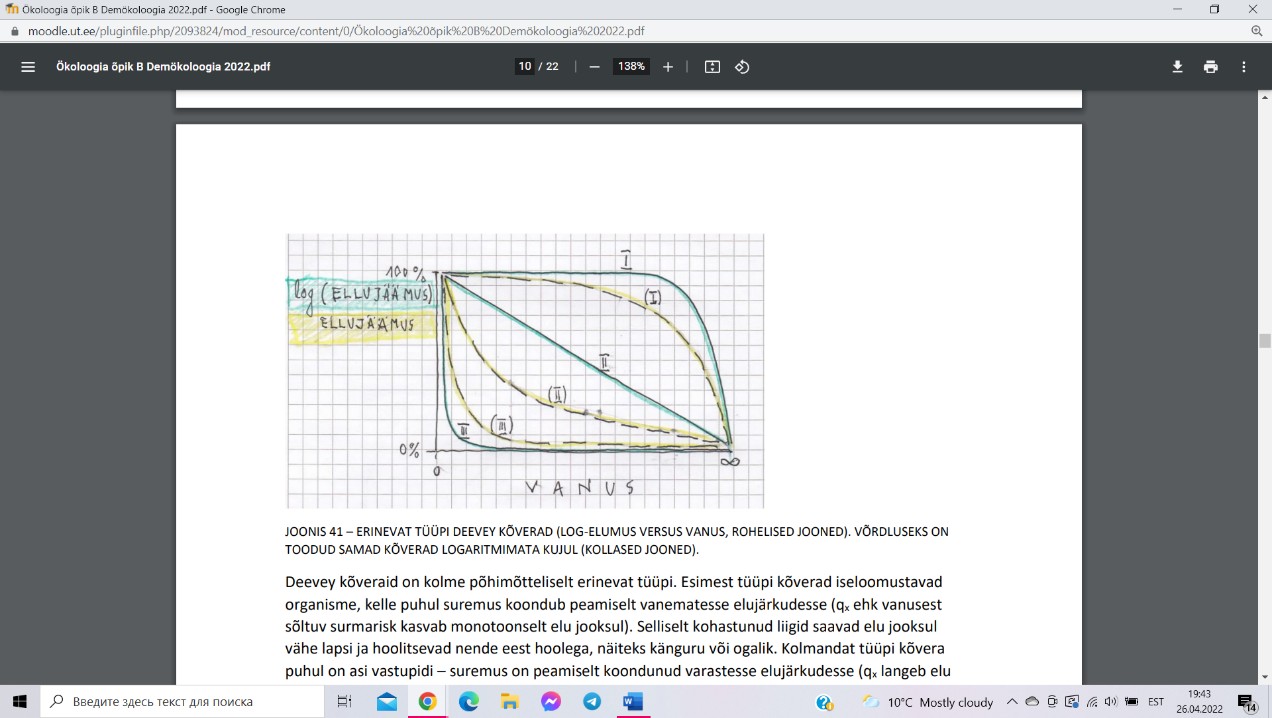
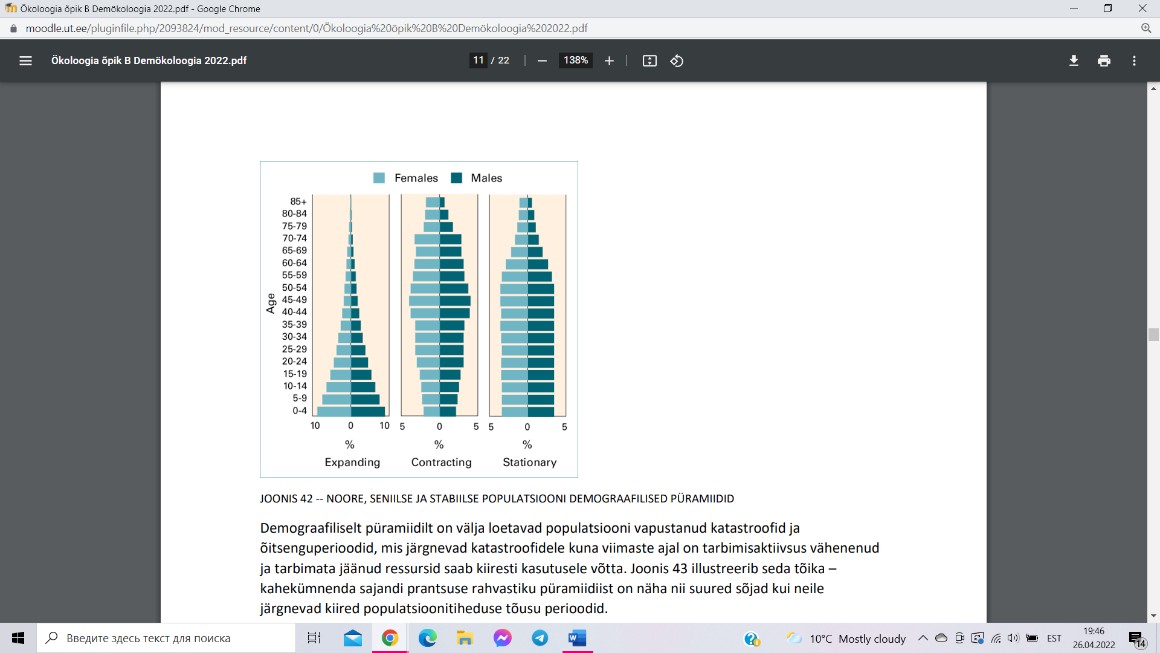
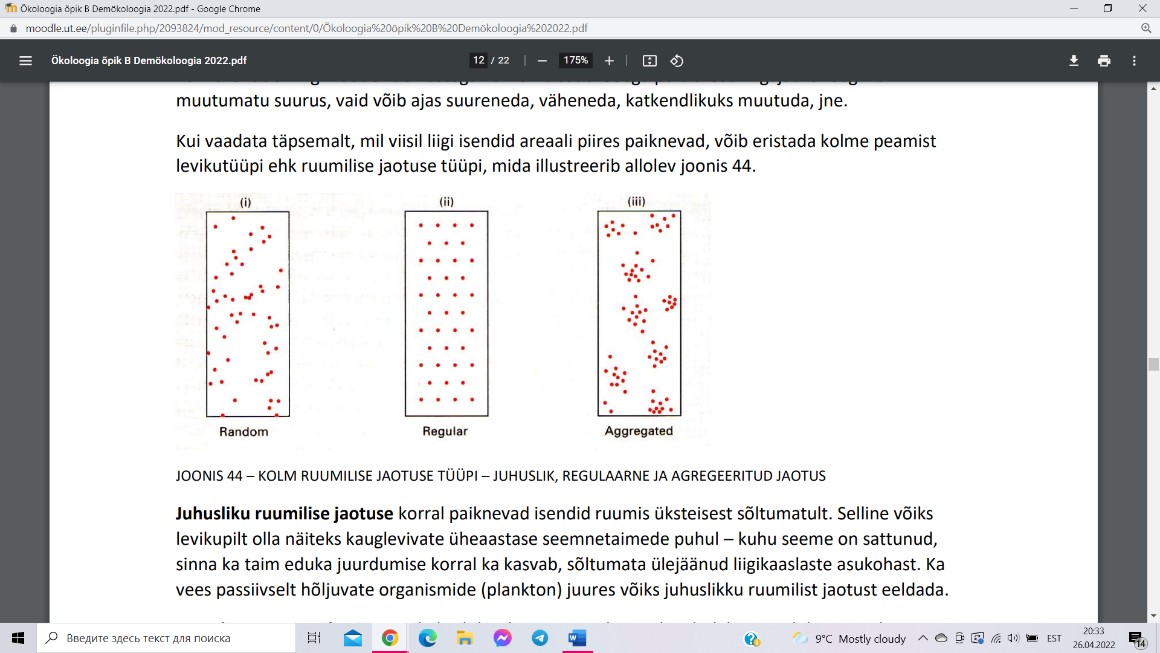
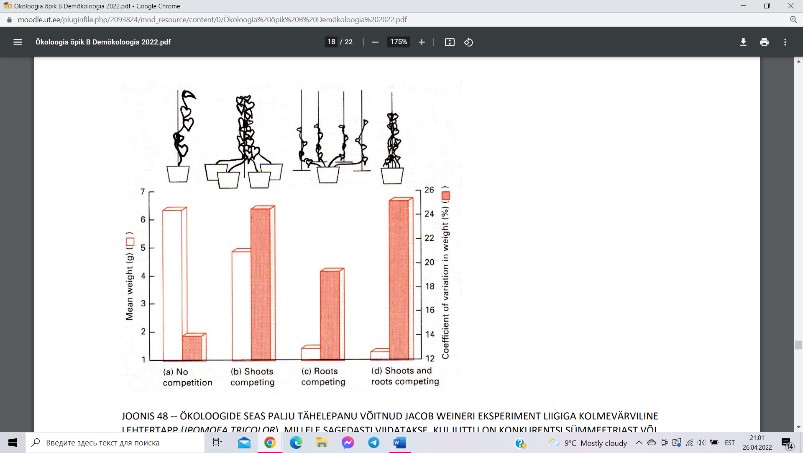
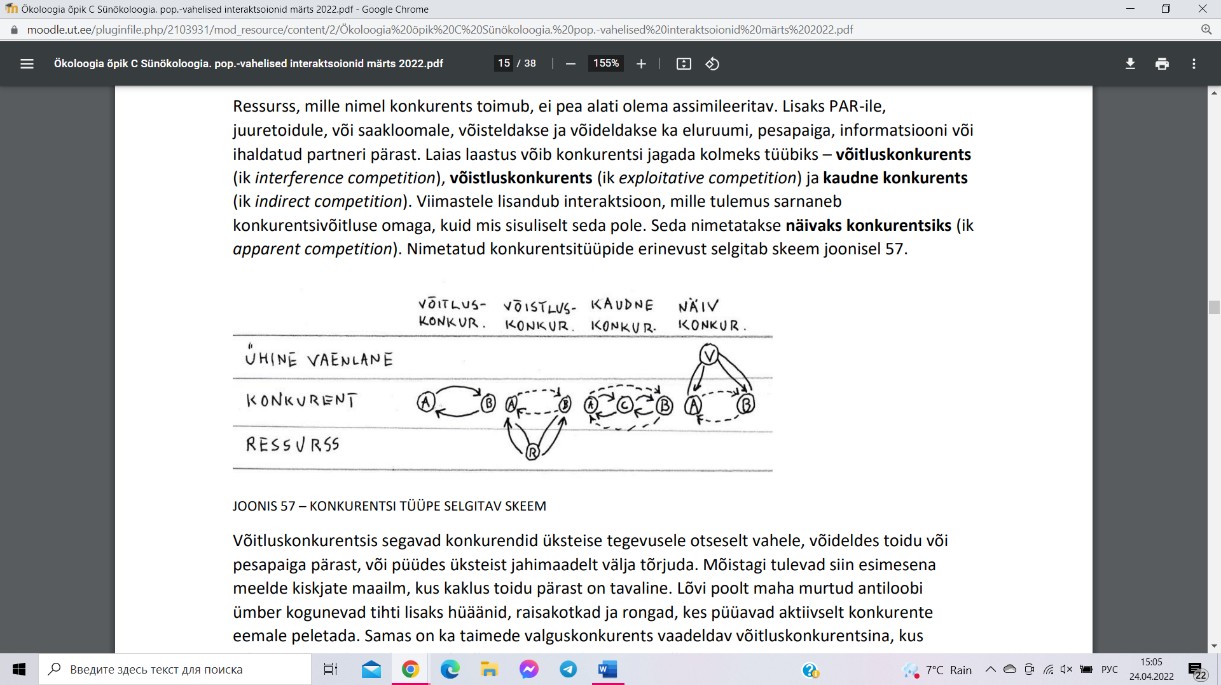
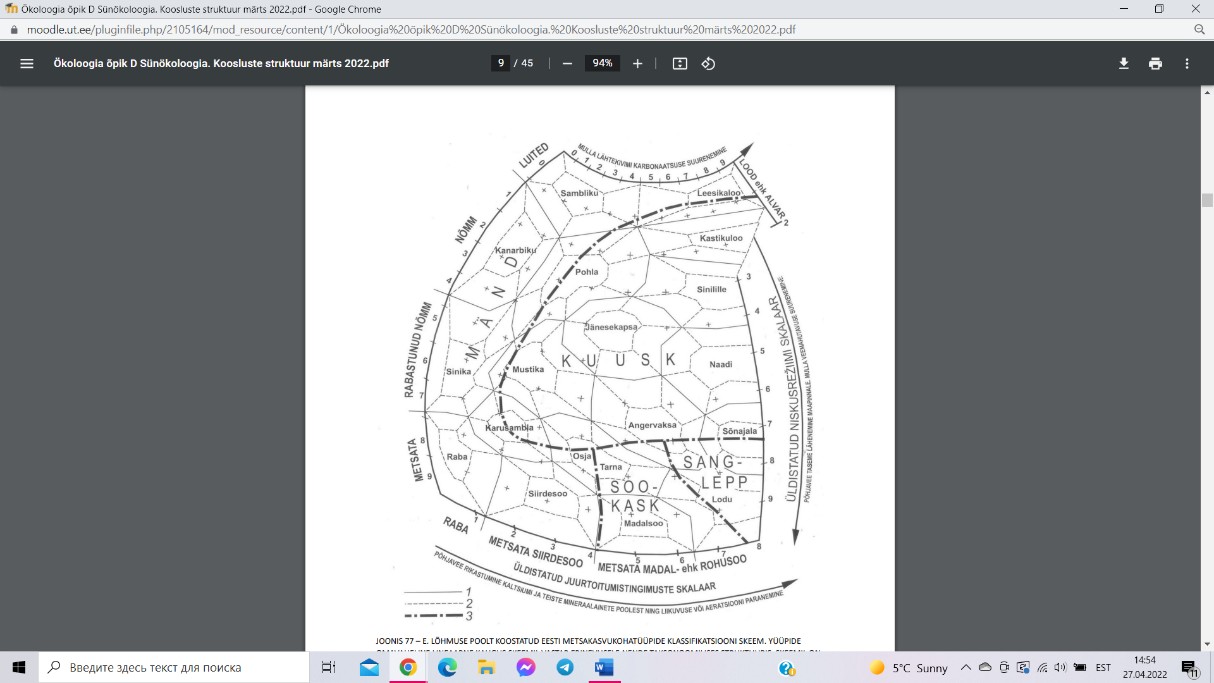
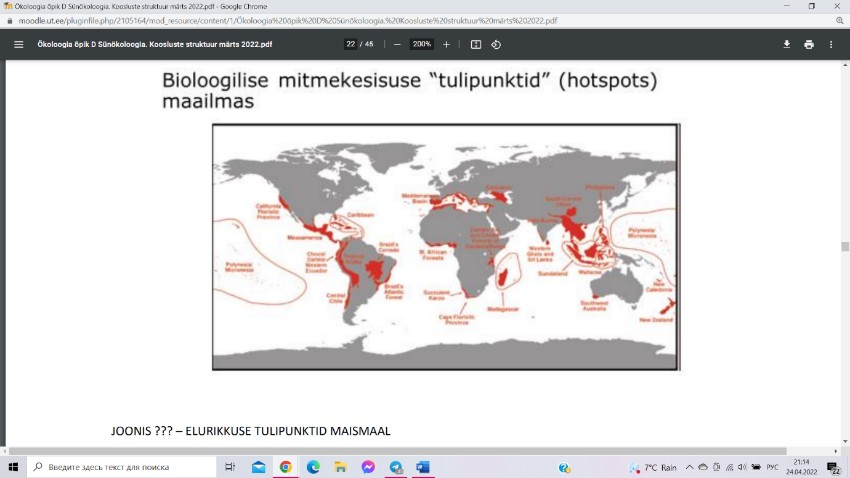
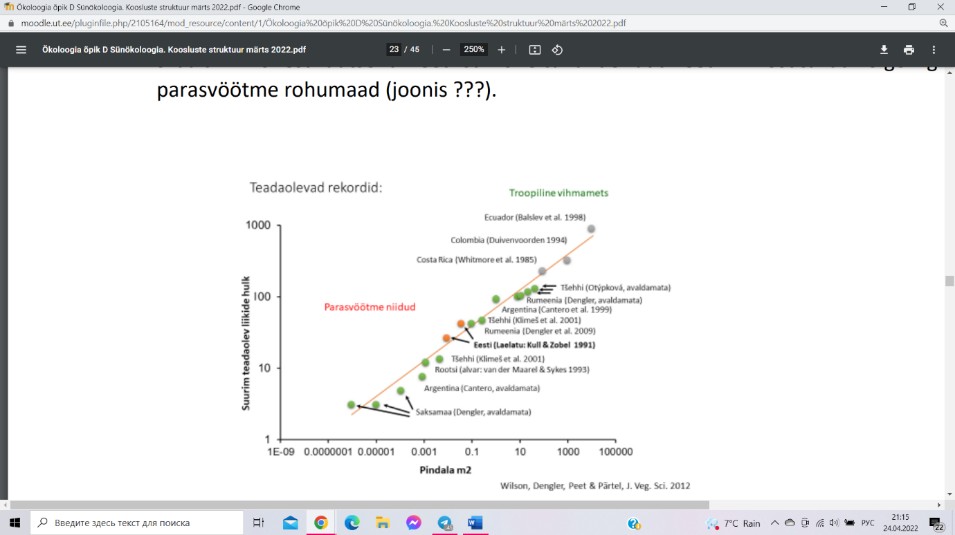
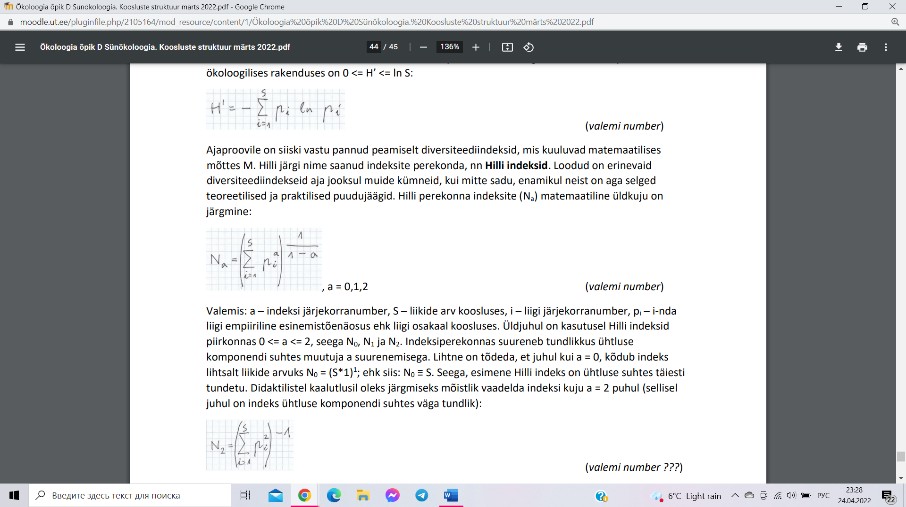
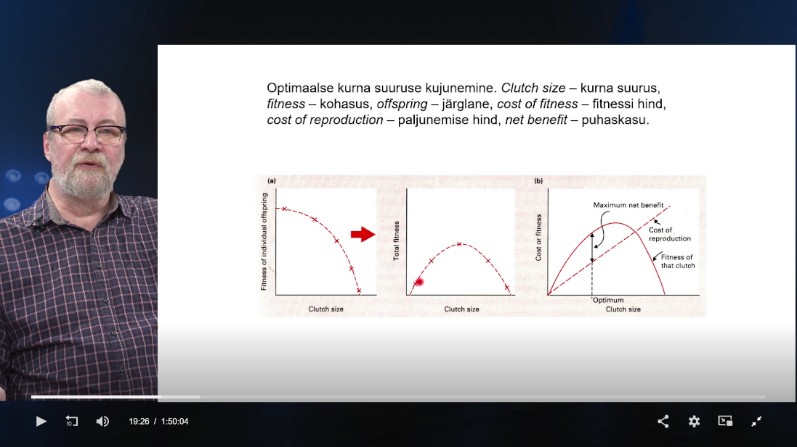

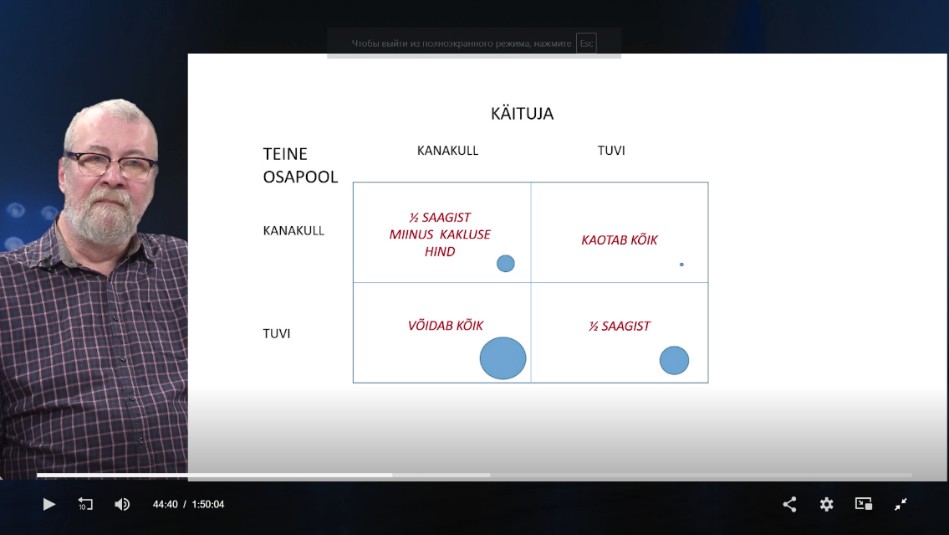

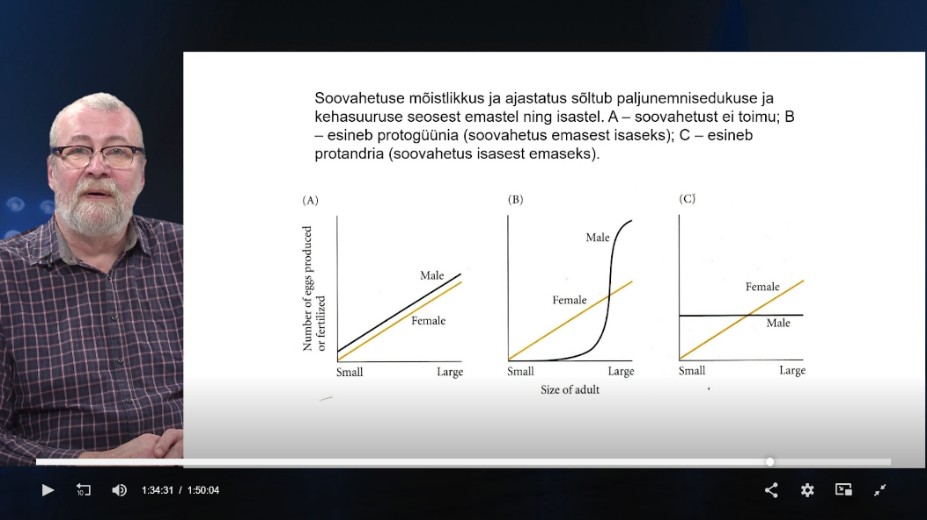
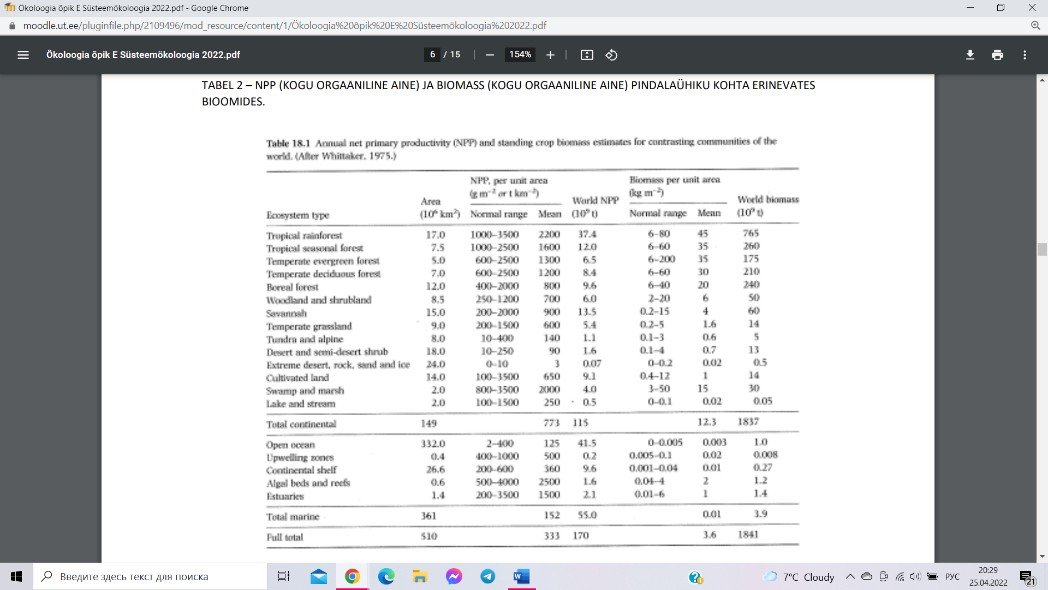
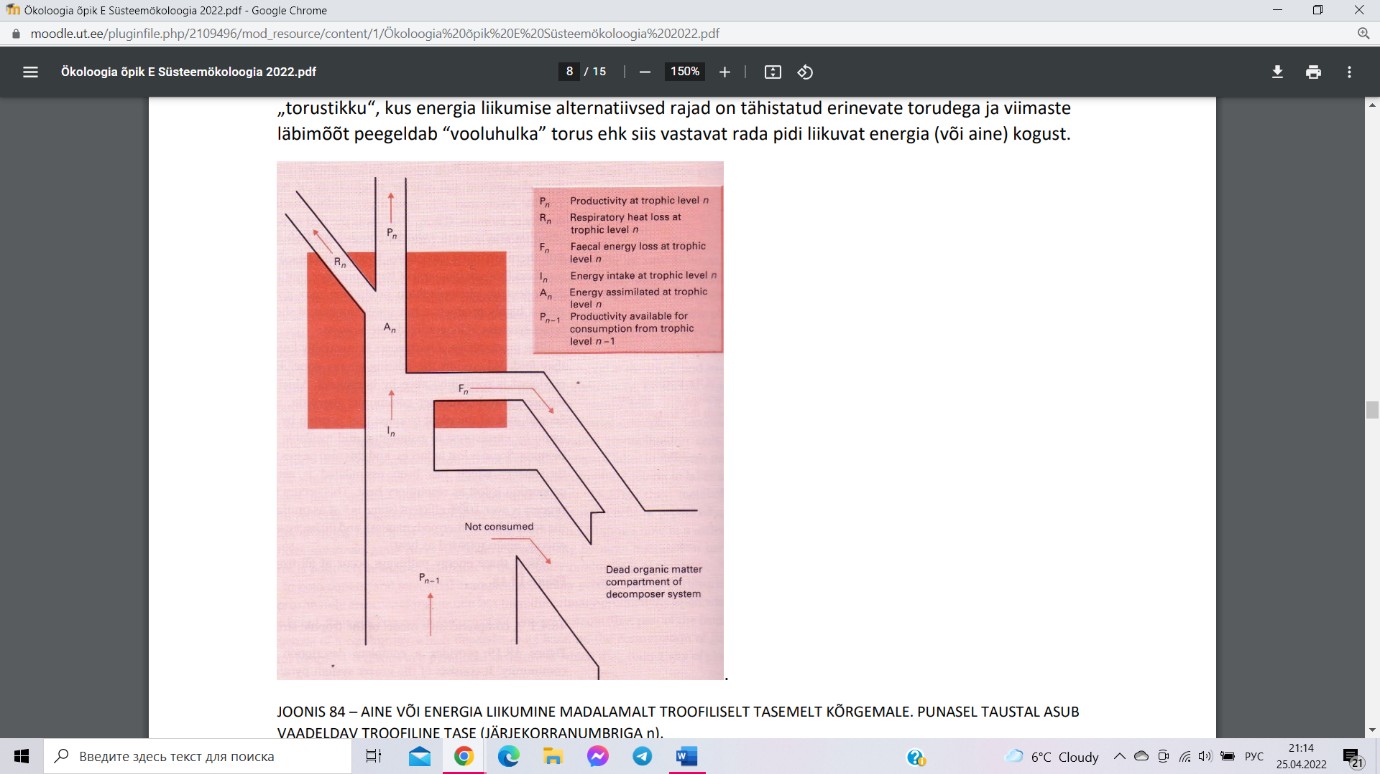
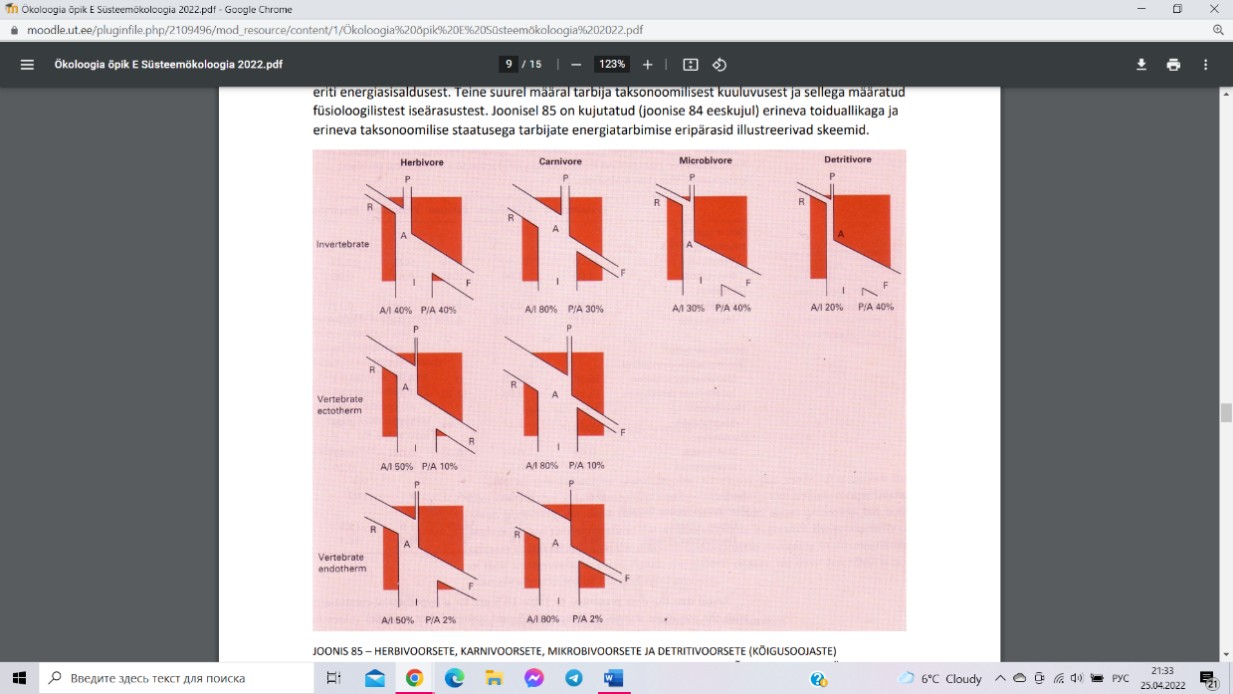
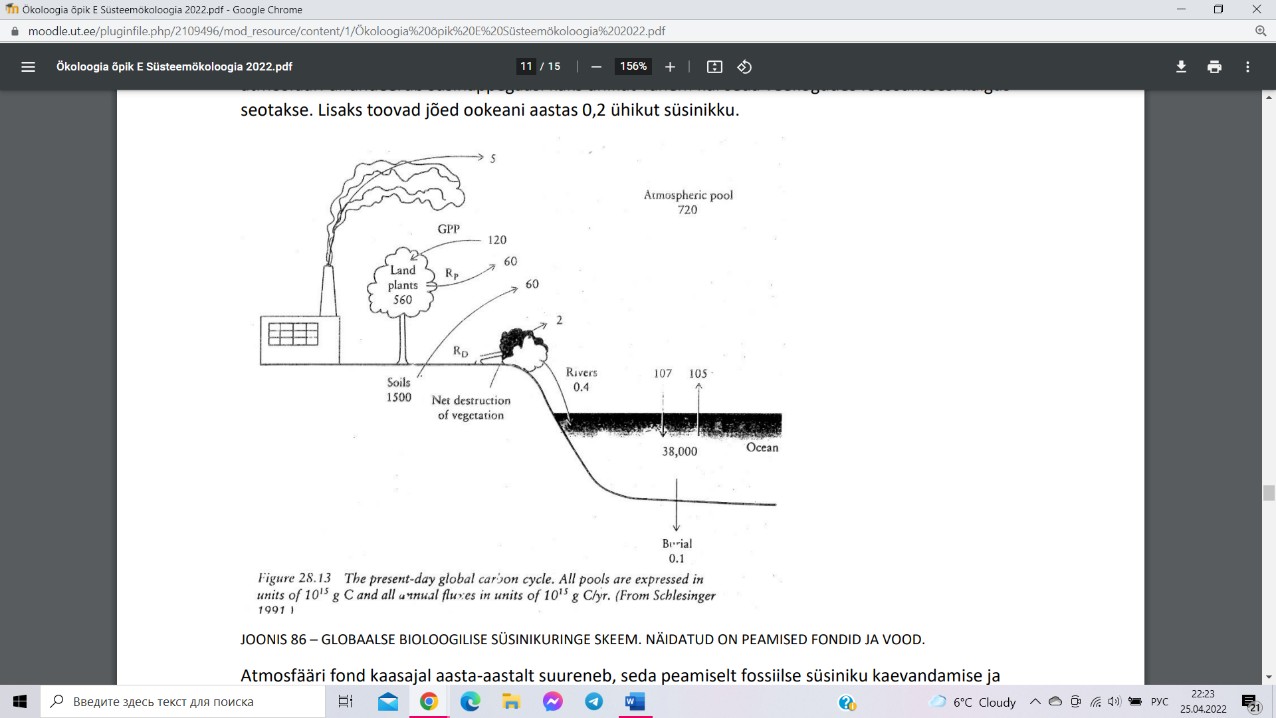
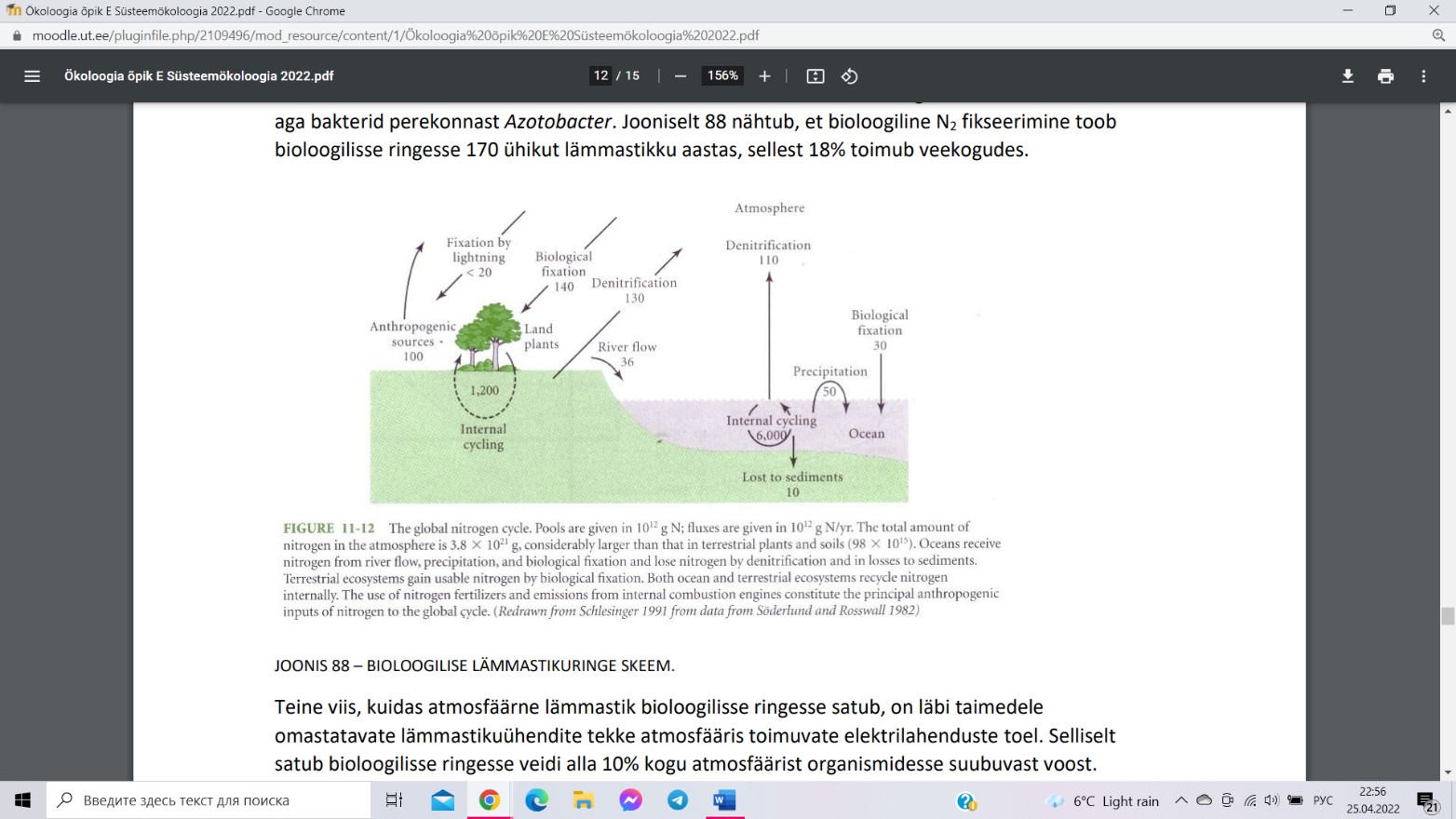
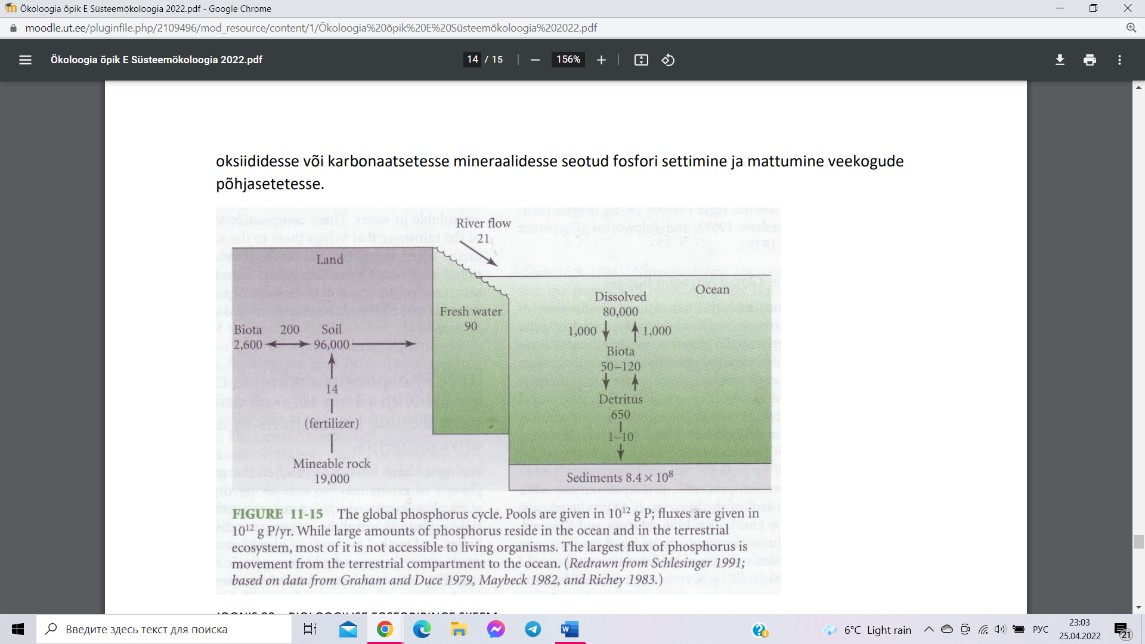
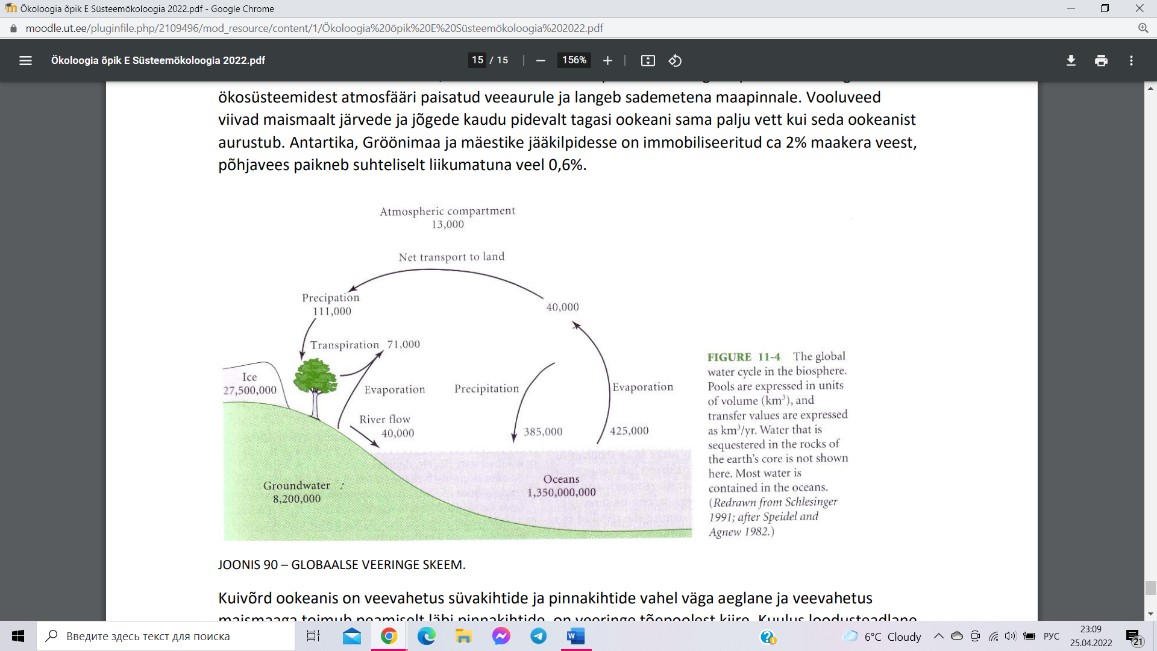
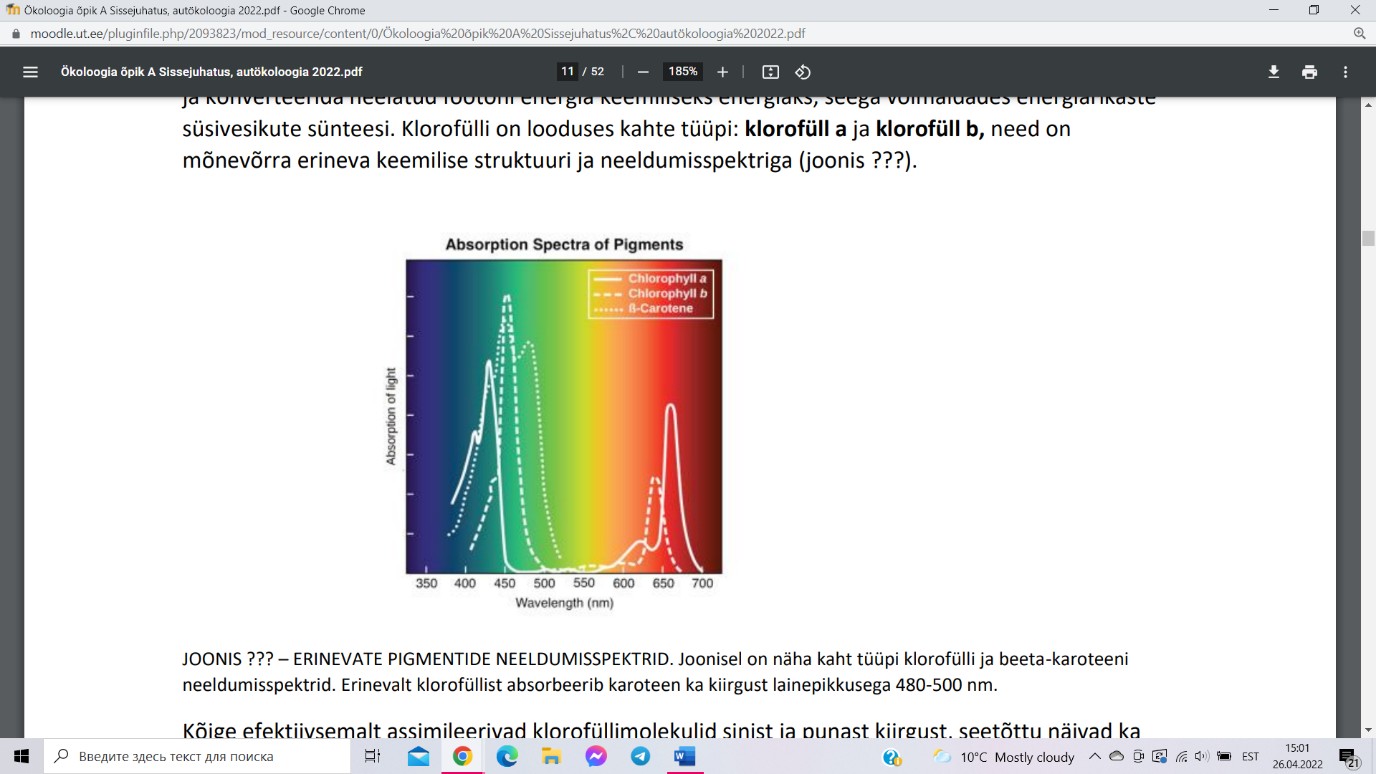
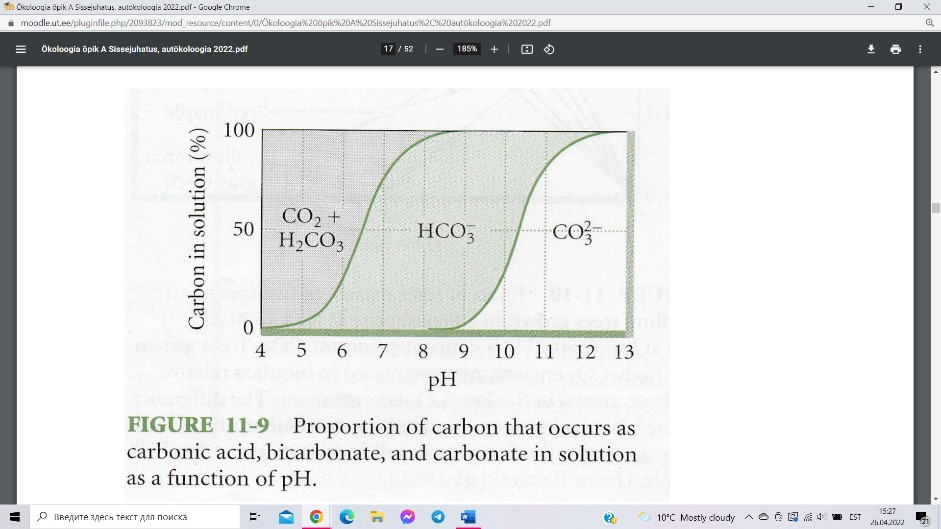
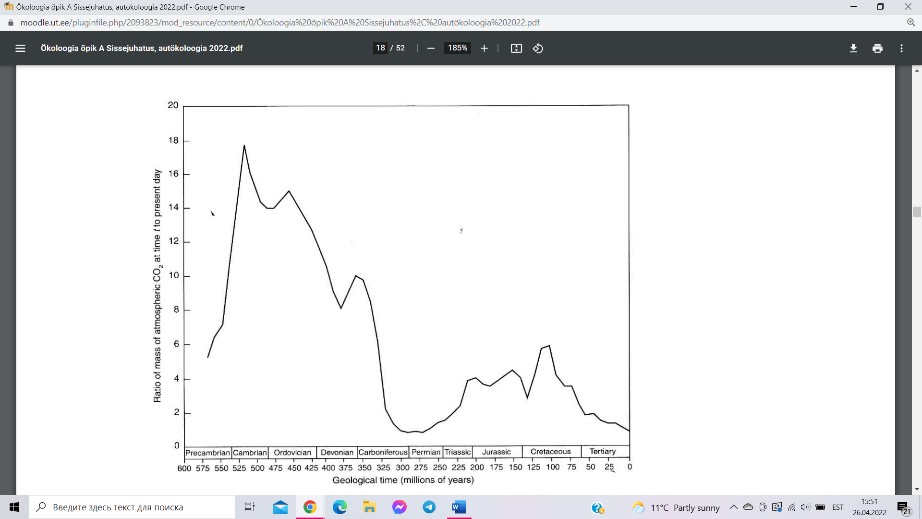
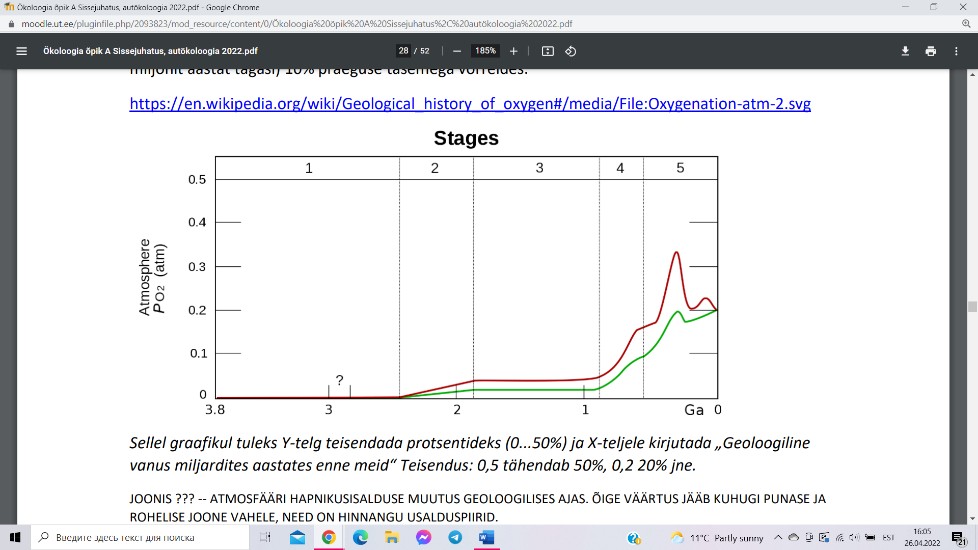
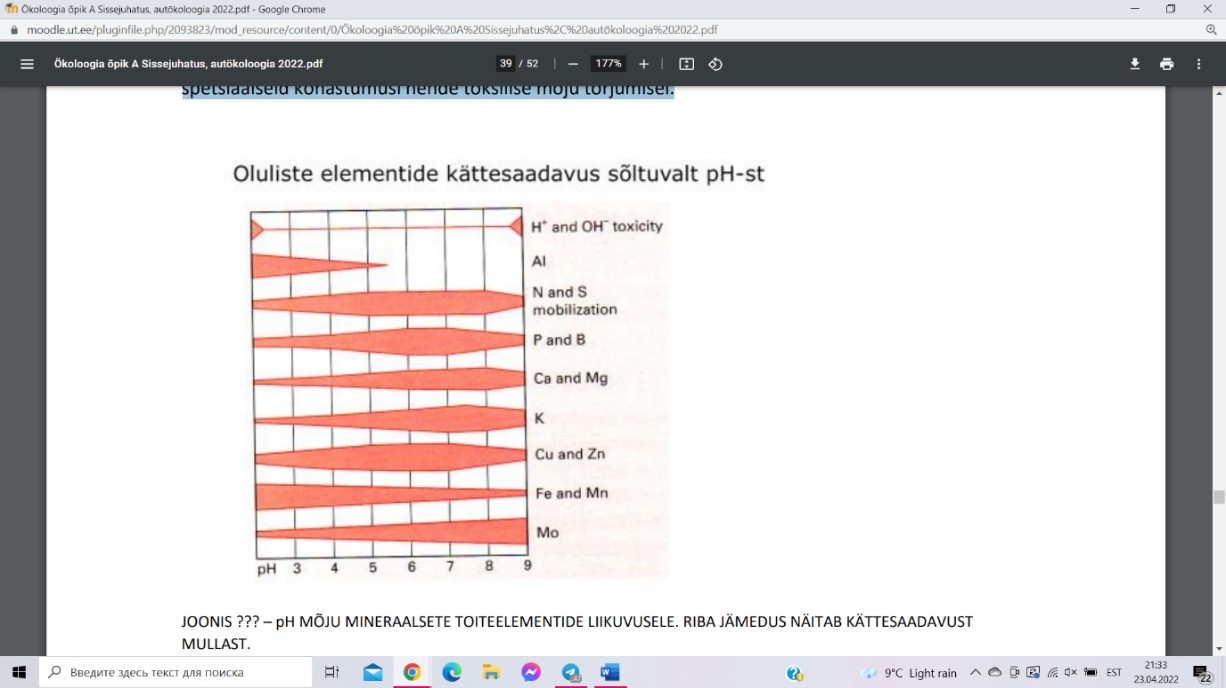
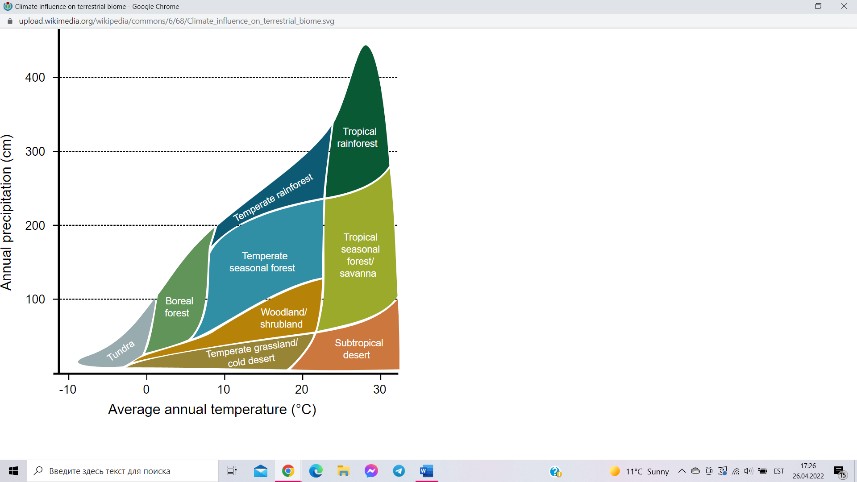
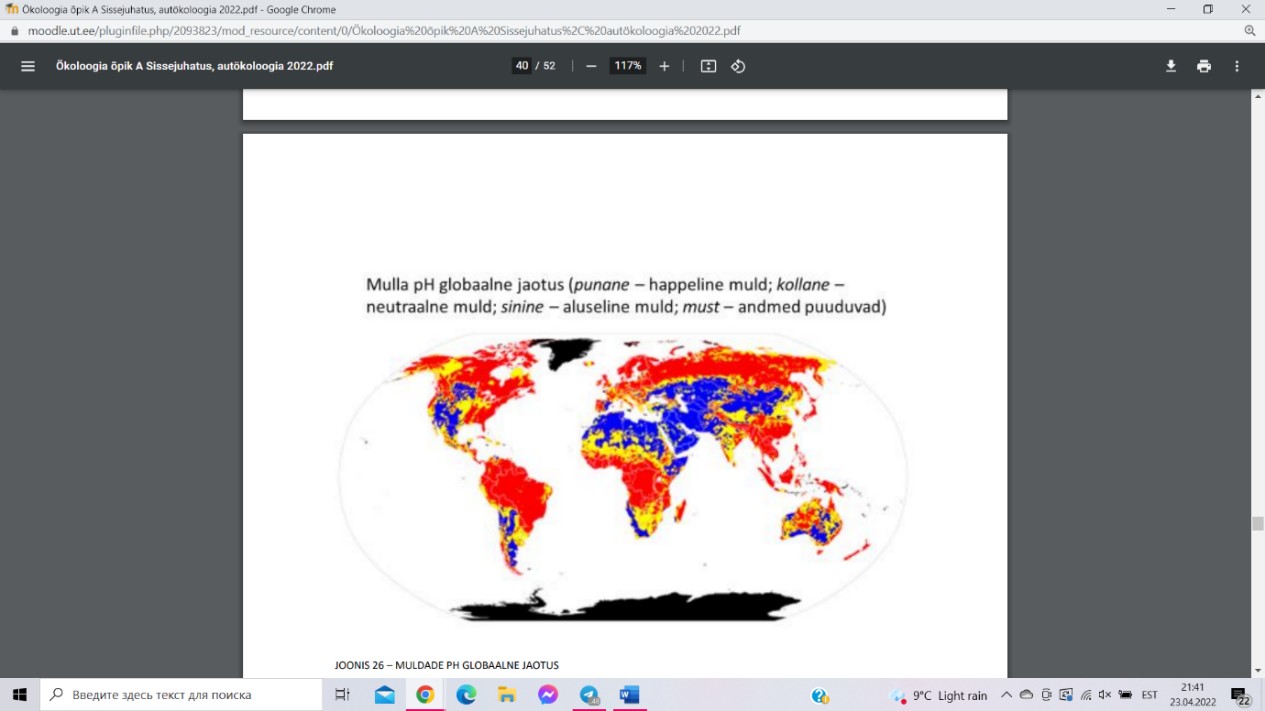
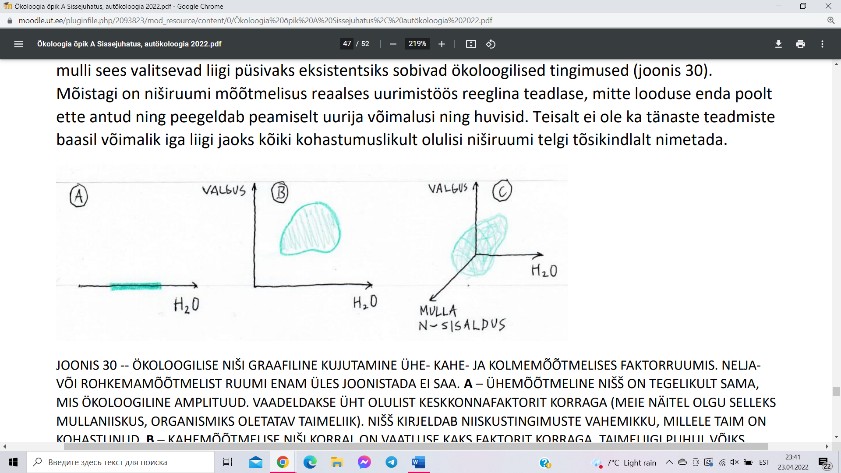
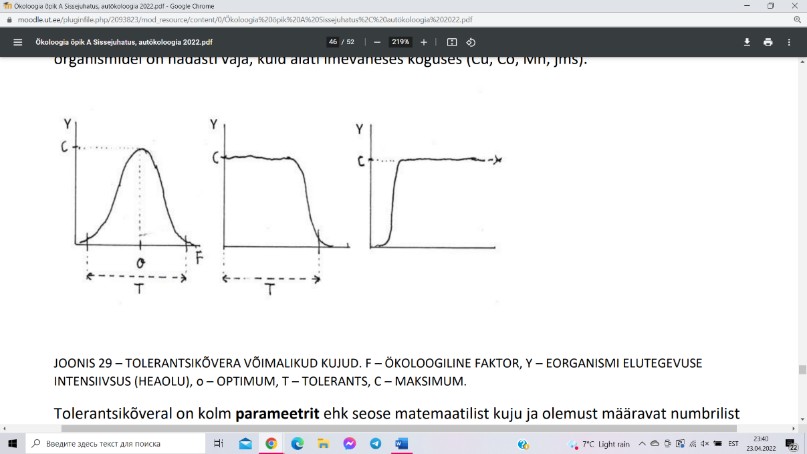
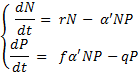
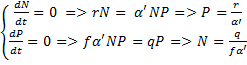










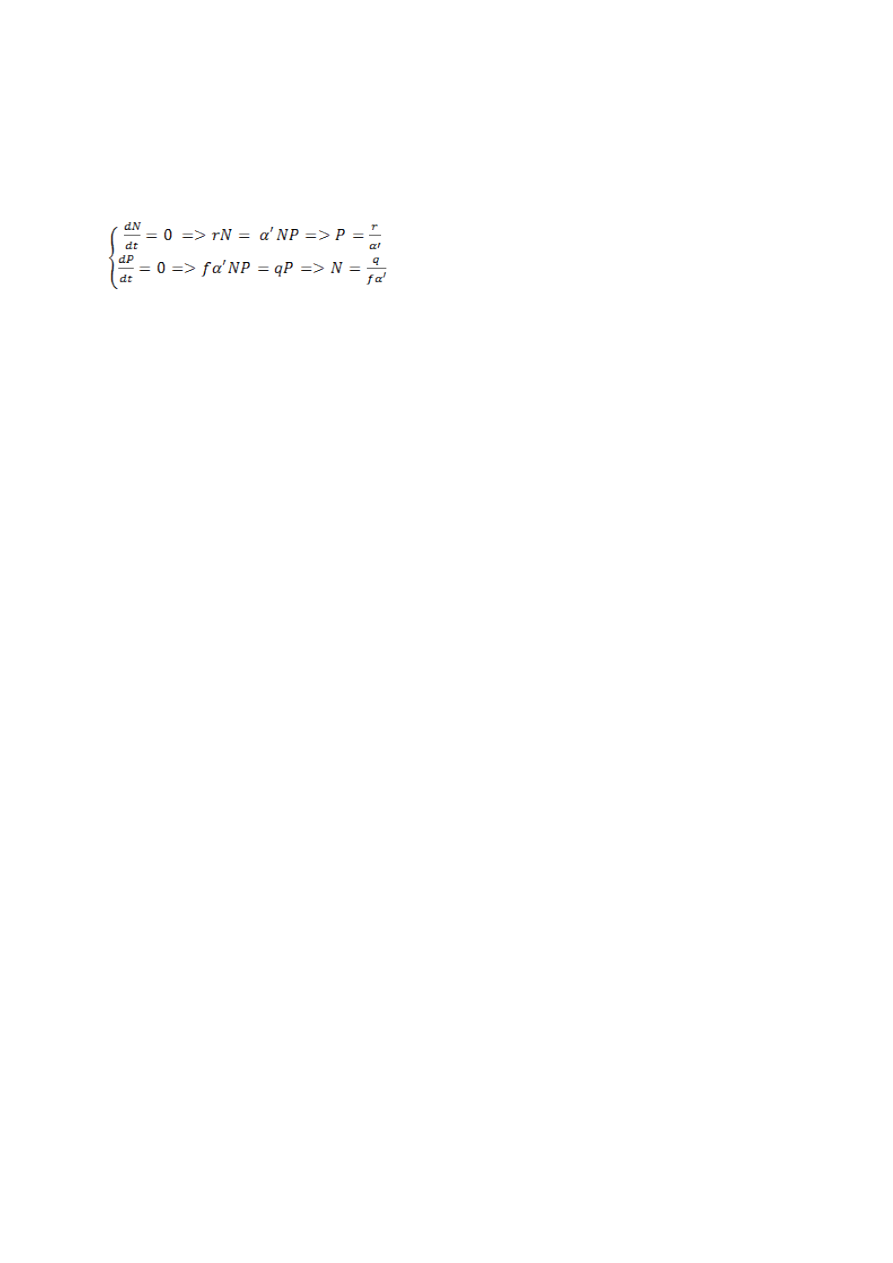

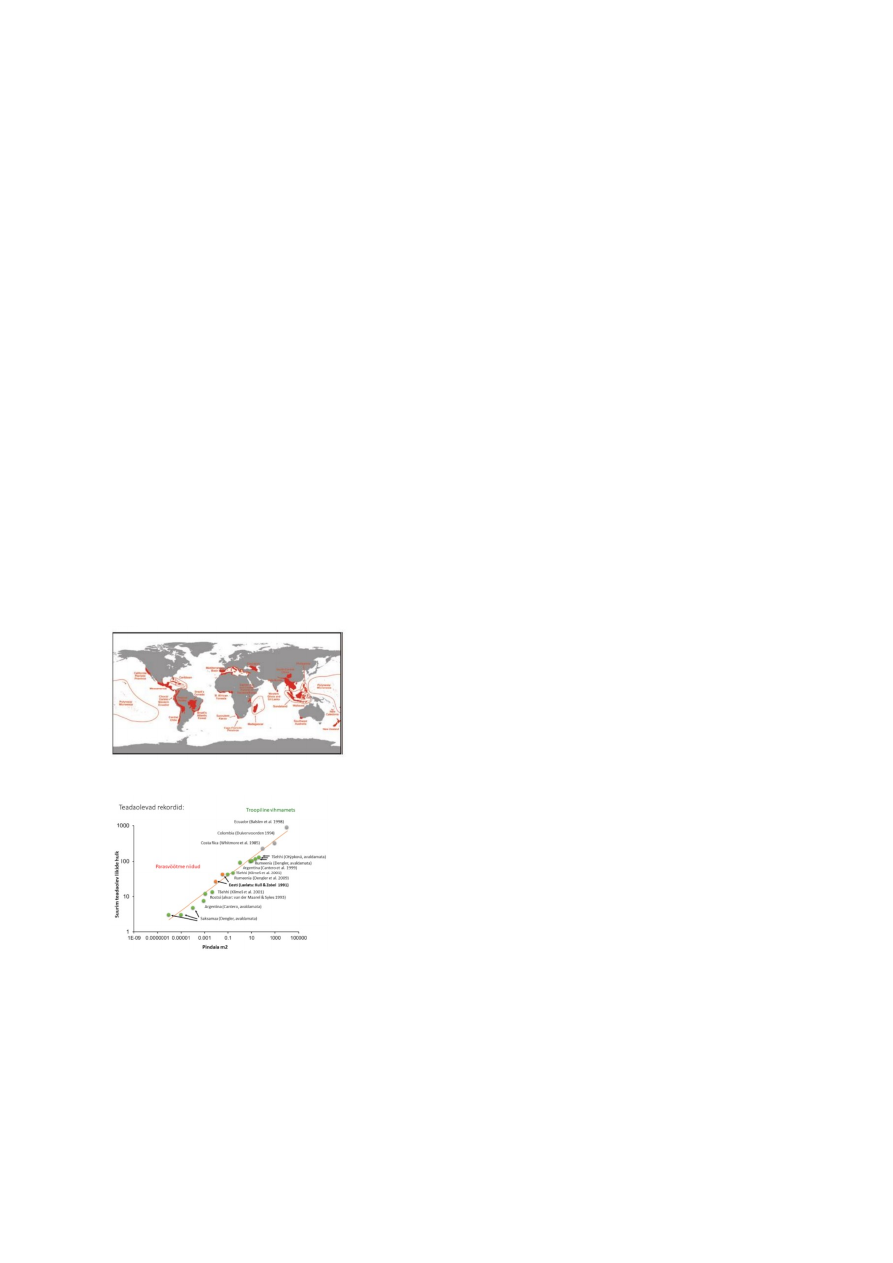




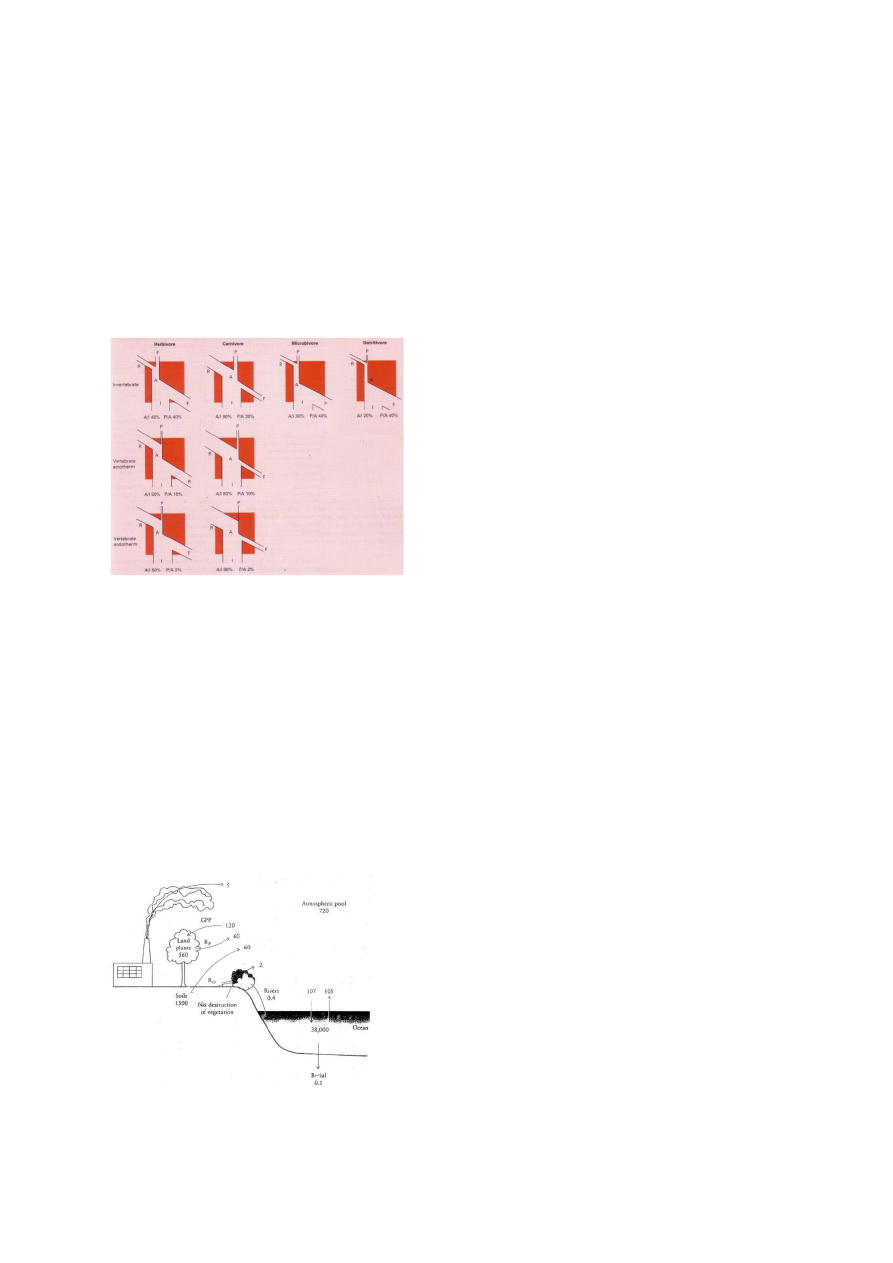
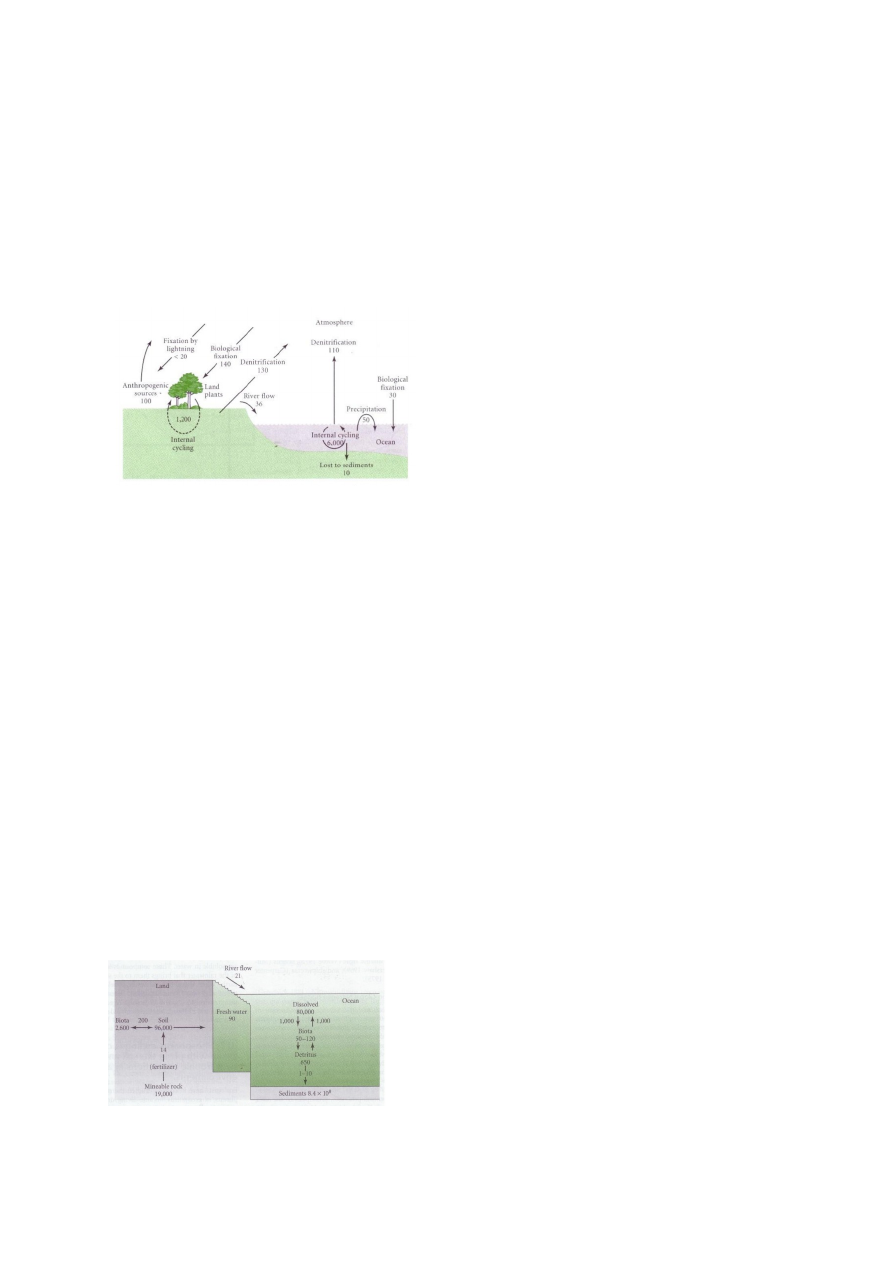





Kõik kommentaarid