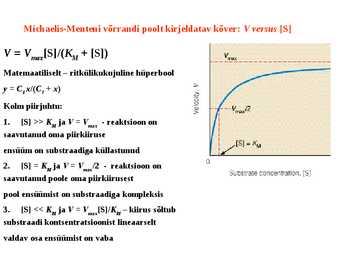
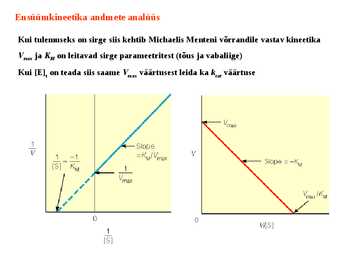
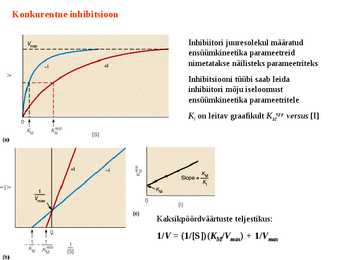
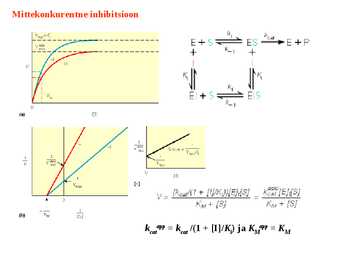
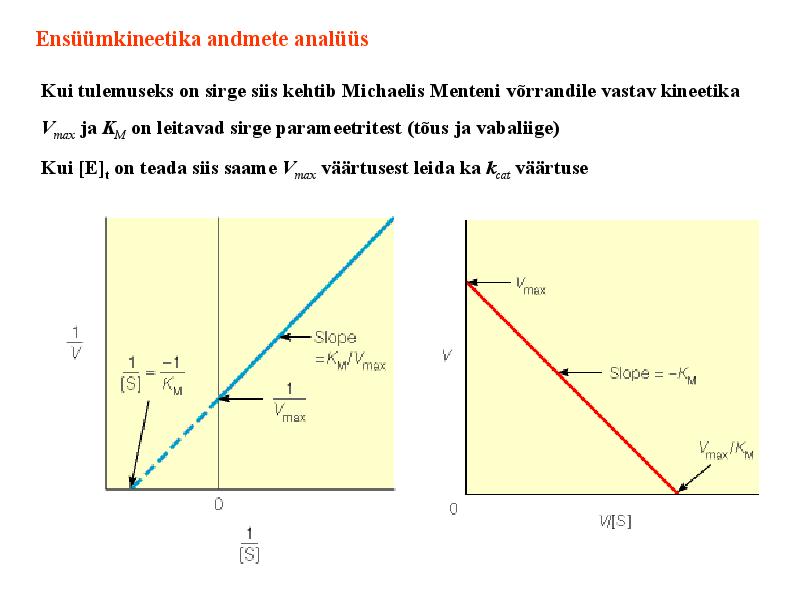
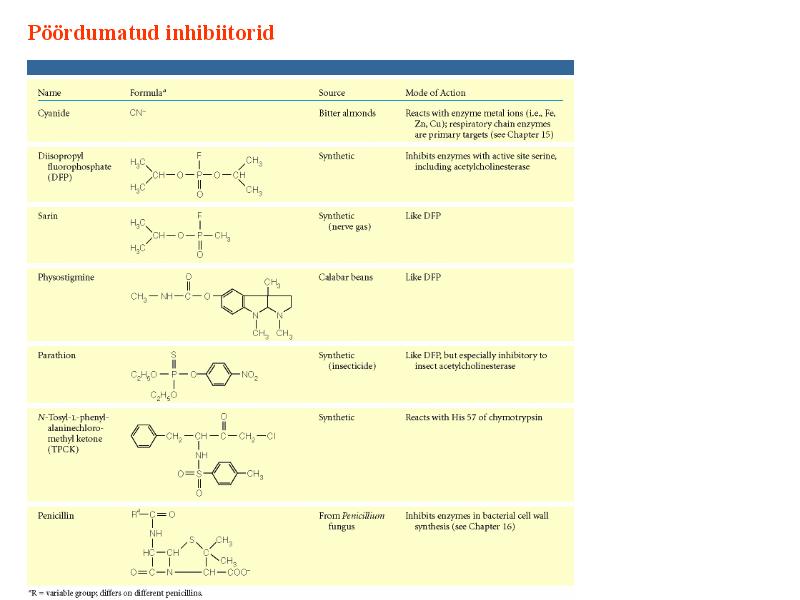
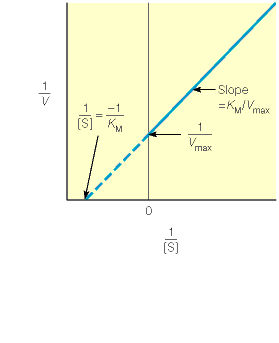
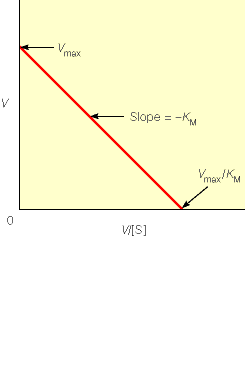
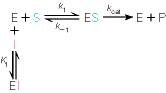
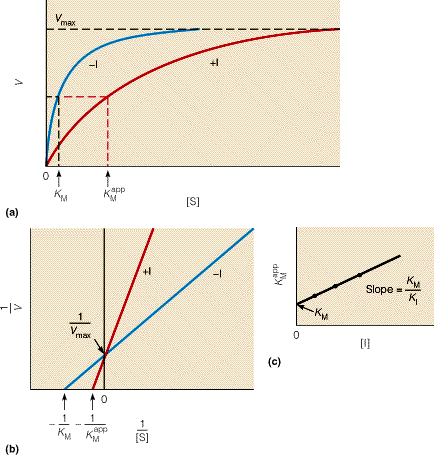
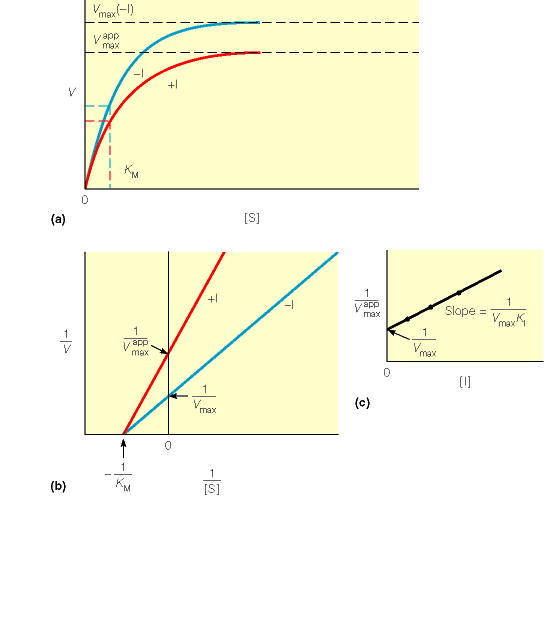
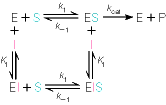
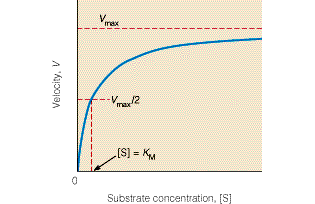
Kuna reaktsiooni substraadi kontsentratsioon on kõrgem, kui ensüümi kontsentratsioon, siis on vaba substraadi kontsentratsioon võrdne kogu substraadi kontsentratsiooniga ja vaba ensüümi kontsentratsioon on võrdne kogu ensüümi kontsentratsiooniga ([E]total), millest tuleb: [E]=[E]t – [ES] ( [ E ]total −[ ES ] )∗[ S ] [ES] ¿ Reaktsiooni max kiirust saavutatakse, kui ensüüm on küllastunud Km substraadiga ([ES]=[E]t), seega Vmax= k2 [E]t 5.4 Kirjutage Michaelis-Menteni võrrand ja selgitage seda graafiku peal. Mida näitavad Km ja Vmax väärtused? Mida näitab kcat? Mida näitab kcat/Km? Km – Michaelise konstant: substraadi kontsentratsioon, mille juures reaktsioonikiirus on ½ maksimaalkiirusest. Seostatakse ensüümi afiinsusega: mida väiksem on K m, seda suurem on
Tuletis on tan . on funktsioonile antud punktis tõmmatud puutuja (sirgjoon) tõusunurga tan. Avaldub kui, Hetkkiirus väheneb antud näites, sest substraati jääb vähemaks. Punase joone tõus on algkiirus, rohelise joone tõus on hetkkiirus 30 min järel, sinise joone tõus on hetkkiirus 1 h järel. Kiirus tee endale hästi selgeks!! Ensüümid mõjutavad reaktsiooni kiirust ainult, ei saa panna midagi tekkima, kui seda ei ole, siis pole, ensüüm ei aita sellisel juhul! 1 Reaktsiooni skeem aA+bBuU+rR. Suured tähed ained ja väikesed stöhhiomeetria kordajad. Biokeemias enamasti need 1 ehk 1 molekul ühes reaktsiooni tsüklis. Pärisuund vasakult paremale, vastasuund paremalt vasakule. v(päri)=reaktsiooni skeem+massitoimeseadus, seega vpärivõrdeline[A]a[B]b, vpäri=kpäri[A]a[B]b. Vaata edasi massitoimeseaduse juurest! vvastas=kvastas[U]u[R]r
ENSÜÜMIKINEETIKA 1. Keemilise kineetika põhimõisted reaktsiooni kiirus aeg, mille jooksul reaktsioon toimub, tähistatakse v tähega. Reaktsiooni kiirus sõltub (enamasti, mõnikord mitte, ntx null-järgu reaktsioon ei sõltu) reageerivate ainete (alg)kontsentratsioonist (ehk kui meil on aine A ja see muutub aineks P, siis kiiruse valem on selline v = d[P]/dt = -d [A]/dt = k [A]). Keemiline kineetika ongi see teadusharu, mis tegeleb reaktsioonide kiiruse uurimisega. kiiruskonstandid kiiruskonstandid on vidinad, mis seovad reaktsioonikiiruse reageerivate ainete kontsentratsioonidega, tähistatakse tähega k (vt ka eelmist mõistet). reaktsiooni järk see näitab, kuidas sõltub reaktsiooni kiirus reageerivate ainete kontsentratsioonidest (kontsentratsioon näitab kui palju ainet ehk selle aine osakesi on meil hetkel
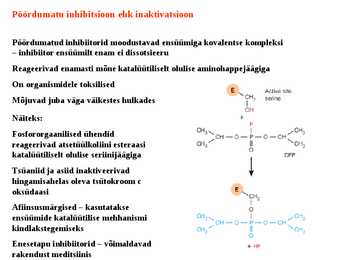
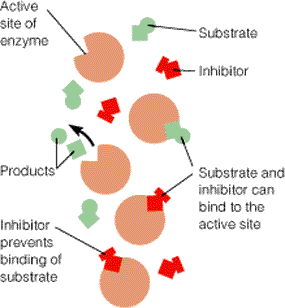
Sellist meetodit kasutavad bakterid just erinevate suhkrute transportimisel rakku. 19. Millised väited on õiged? Katalüsaator: Katalüsaator kiirendab keemiliste reaktsioonide toimumist ja kiirendab spetsiifiliselt just pärisuunalist reaktsiooni. 20. Katalaas katalüüsib vesinikperoksiidi lagundamist. Kas katalaasi hulk reaktsiooni käigus: Katalaasi hulk vesinikperoksiidi lagundamisel ei muutu, sest katalaas on katalüsaator ja katalüsaatori hulk reaktsiooni käigus ei muutu. 21. Ensüüm katalüüsib pöörduvat reaktsiooni A B. Mis hakkab toimuma ensüümi lisamisel aine B lahusele? Hakkab moodustuma ainet A, sest katalüsaator suurendab eelkõige pärisuunas toimuvat reaktsiooni. 22. Vaatame pöördumatut reaktsiooni A B. Kuidas avaldub reaktsiooni kiirus aine A kontsentratsiooni kaudu? V= 23. Mitmendat järku reaktsiooniga on tegemist? (erinevad reaktsioonid) a) E + L EL 2 järku (E)+(L) b) EL E +L 1 järku (EL) 24
Regulatsioon Rakk on piltlikult võrreldav tööstusettevõttega · Vabrik Rakk · Toorained Lähteained · Valmistoodang Lõpp-produktid · Vaheproduktid metabolismi vaheühendid · Tööpink Ensüüm · Tootmisliin Metabolismirada Vabriku töö peab olema reguleeritud: erinevate tööpinkide läbilaskevõime koordineerimine erinevate tootmisliinide läbilaskevõime koordineerimine tooraine jaotus erinevate tootmisliinide vahel turu nõudlus erinevate valmistoodete järele Vabrikus on insenerid ja ülemused Rakkudes toimib rida erinevaid regulatsioonimehhanisme Reguleeritakse ensüümkatalüüsitavate reaktsioonide toimumise kiiruseid:
Ensümoloogia alused. Kordamisküsimused Ensüüm kui valk: valgu struktuur, aminohapped, mittekovalentsed interaktsioonid, vesilahused ja unikaalsed vee omadused. Valgu funktsioneerimise tagab tema struktuur. Ensüüm kui katalüsaator: keemiline reaktsioon, termodünaamika, kineetika, katalüüs, mehhanism, ensüümide kasutamine tööstuses. Ensüüm kui bioloogiline katalüsaator: sidustatud reaktsioonid, bioenergeetika, metabolism, regulatsioon, klassifikatsioon ja nomenklatuur. Ensüümid on organismide tööhobused. 1) Ensüümkatalüüsi põhimõisted ja printsiibid + Ensüümkatalüüsi peamised tunnus- jooned. · Ensüümkatalüüs põhineb rangelt füüsikalistel ja keemilistel vastasmõjudel. · Kõik ensüümid on evolutsioonilise arengu produktid ja kujunenud selliseks, nagu me
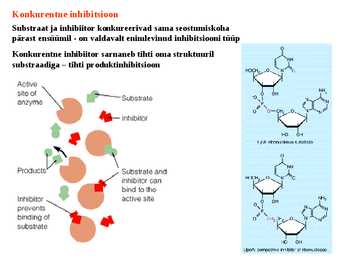
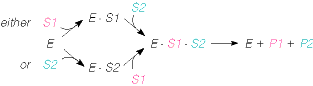
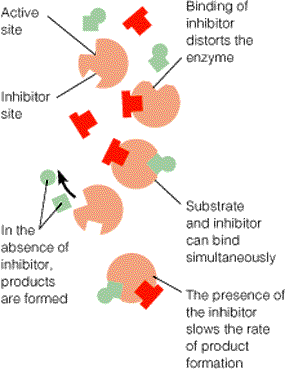
Number (1) fosfotransferaasid, kus fosforüülrühma aktseptoriks on D-glükoosi hüdroksüül- rühm Triviaalnimetus: heksokinaas 3. ES kompleks ja selle formeerumist kirjeldavad molekulaarsed mudelid. E ja S vahelised interaktsioonid. E + S ES P + E ensüüm substraat ensüümi- produkt ensüüm substraadikompleks Substraat seotakse ensüümile nõrkade jõudude toimel vesiniksidemed, van der waalsi interaktsioonid, ioonsed sidemed, mõnel korral ka hüdrofoobsed interaktsioonid. ES kompleksi moodustamisega kaasneb entroopia vähenemine ES kompleksi moodustamisel toimub substraadi desolvatatsioon LIISI KINK 23
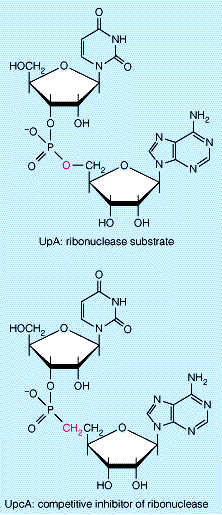
kristalliseeruvad · kõrgspetsiifilised tuleneb valgulisest loomusest ja see läbi ensüümi aktiivtsentri ja substraadi komplementaarsusest. See määrab suuresti protsesside kulu vajalikus suunas ja koordineerituses. - substraadispetsiifilisus- spetsiifilisus ühe substraadi suhtes (nt lipaasid lõhustavad ainult lipiide). *absoluutne spetsiifilisus ensüüm muundab vaid ühte substraati (arginaas muundab Arg, mitte metüülArg) *stereokeemiline spetsif E muundab vaid ühte stereoisomeeri (L-aminohappe oksüdaas toimib vaid L- aminohappele) *sidemespetsif sahharaas lõhustab vaid glükoosi ja fruktoosi vahelist glükosiidsidet sahharoosis *rühmaspetsiifilisus nii pepsiin kui ka trüpsiin hõdrolüüsivad peptiidsidemeid




































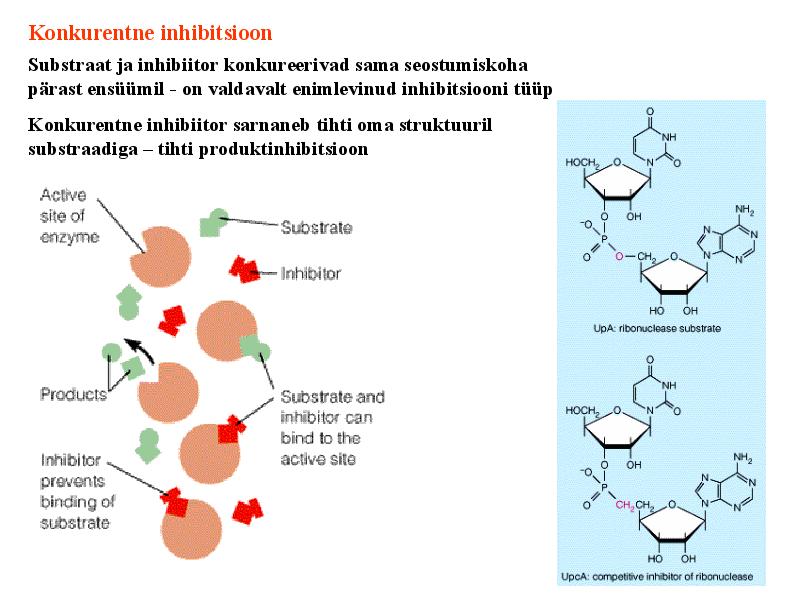

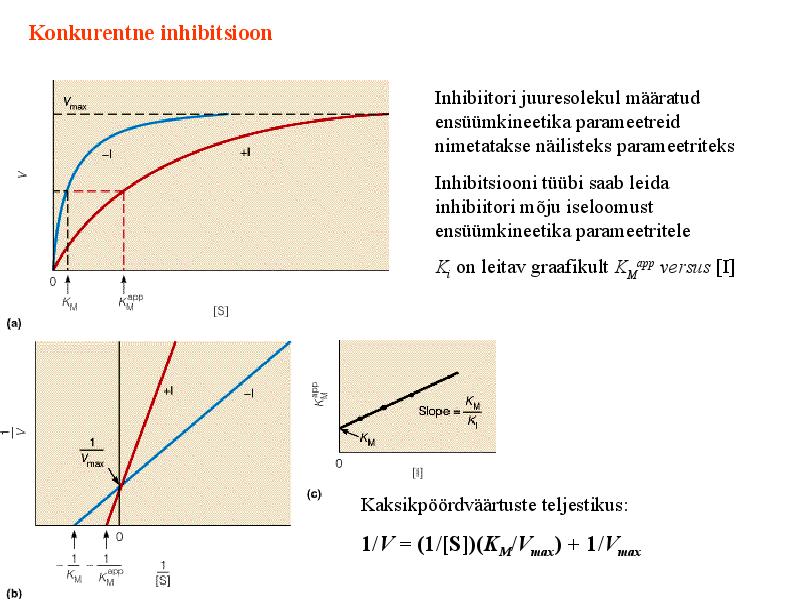


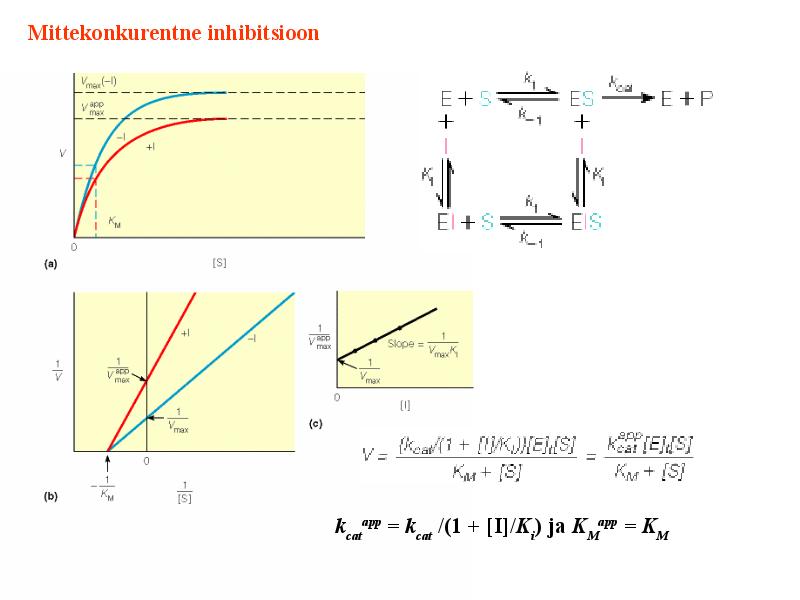
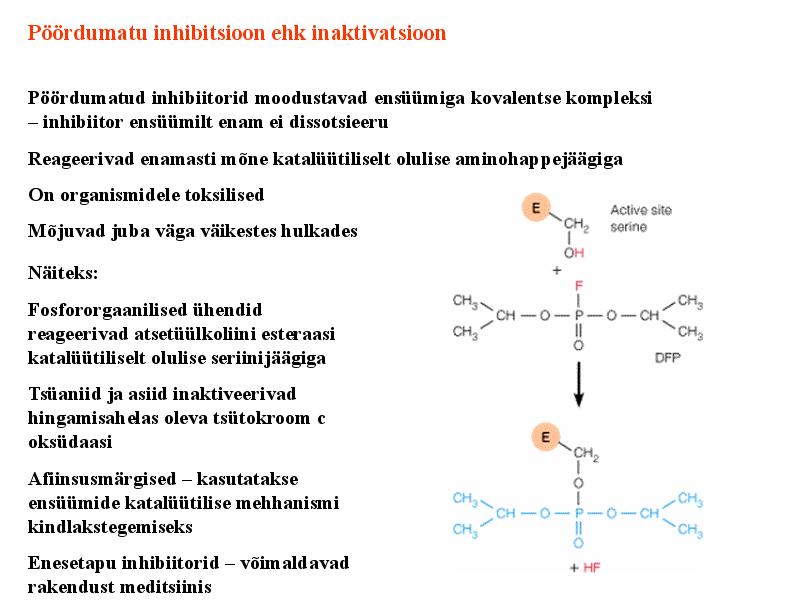








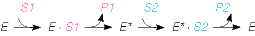

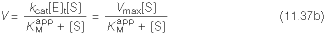
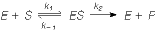





Kõik kommentaarid