74. Millised on need kaks RNA omadust, mis annavad alust pidada just RNA-d keskseks molekuliks elu varases evolutsioonis? Kaks RNA omadust, mis annavad alust pidada just RNAd keskseks molekuliks elu varases evolutsioonis on potentsiaalne isereplitseerumise võime ja võimalik katalüütiline aktiivsus. 75. Mis on metabolismi regulatsiooni lõplikuks eesmärgiks? Metabolismi regulatsiooni lõplikuks eesmärgiks on biokeemiliste reaktsioonide toimumise kiiruse kontroll. 76. Kuidas toimib regulatsioon substraadi tasandil? Reaktsioonide kiiruste reguleerimine on saavutatav muutuste kaudu substraatide ja produktide kontsentratsioonis. 77. Ensüümi KM substraadi jaoks on 10 mM. Substraadi kontsentratsioon rakus on: a) 1,0 mM b) 10 mM c) 100 mM Millisel juhul peegeldub substraadi kontsentratsiooni 20% -line kasv kõige paremini reaktsioonikiiruse kasvus, millisel juhul kõige halvemini? (võivad olla erinevad arvud) 78. Substraadi kontsentratsioon rakus kasvab 20 M -lt 40 M-ni
muundavadki S-di. · Liitensüümides on aktiivtsentris ko koensüüm või muu kofaktor · substraat fikseeritakse katalüüsiks vajalikku asendisse paljude sidemetega · aktiivtsentri lõplik ruumiline formeerumine toimub substraadimoekuli lähenemisel: S ja E komplementarsuse alusel muudetakse aktiivtsenter S-le sobivaks isosteeriline regulatsioon substraadi poolt. · S ja E interaktsioonil muutub ensüümi konformatsioon: S-di ja aktiivtsentri katalüütiliste rühmade kontakt muutub sobivamaks, ,,pingustavad" sidemed substraadis. Seetõttu on ensüüm-substraat kompleksi (ES) teke ja substraadi muundumine produktides kergendatud. · Tekkiv produkt pole komplementaarne aktiivtsentri katalüütilise rühmadega, seetõttu ta väljub aktiivtsentrist ja selle algkonformatsioon taastub. Ensümaatiline kineetika
Ensümoloogia alused. Kordamisküsimused Ensüüm kui valk: valgu struktuur, aminohapped, mittekovalentsed interaktsioonid, vesilahused ja unikaalsed vee omadused. Valgu funktsioneerimise tagab tema struktuur. Ensüüm kui katalüsaator: keemiline reaktsioon, termodünaamika, kineetika, katalüüs, mehhanism, ensüümide kasutamine tööstuses. Ensüüm kui bioloogiline katalüsaator: sidustatud reaktsioonid, bioenergeetika, metabolism, regulatsioon, klassifikatsioon ja nomenklatuur. Ensüümid on organismide tööhobused. 1) Ensüümkatalüüsi põhimõisted ja printsiibid + Ensüümkatalüüsi peamised tunnus- jooned. · Ensüümkatalüüs põhineb rangelt füüsikalistel ja keemilistel vastasmõjudel. · Kõik ensüümid on evolutsioonilise arengu produktid ja kujunenud selliseks, nagu me neid täna näeme, evolutsiooni ja loodusliku valiku tulemusel.
ENSÜMOLOOGIA Lp tudengid. See konspekt on kirjutatud tudengite, kelle nimed on mulle paraku teadmata, poolt. 2013 aastal täiendas konspekti magistrant Karl Annusver, kes lisas joonised ja tegi võrrandid paremini jälgitavaks. Konspekt on kirjutatud seotult loengus näidatavate slaididega. Konspekt on minu poolt läbi vaadatud ja suuremaid möödalaskmisi ei sisalda. Päris iseseisvaks õppimiseks see siiski mõeldud ei ole. Edukat ensümoloogia õppimist ja tänud anonüümsetele autoritele ning Karl Annusverile! Priit Väljamäe 20.11.2017 ,,Structure and mechanism on protein science" Alan Fersht Biokeemia põhiõpik, kus ensümoloogia ka sees. Ensüüm keemiliste reaktsioonide katalüsaator (kiirendaja). Iseloom molekulina pole oluline, struktuur pole samuti. Vaatame ainult, mida ta teeb! Substants, mis kiirendab keemiliste reaktsioonide toimumist on katalüsaator. Ise jääb reaktsiooni lõppedes muutumatule kujule. Keemilisele reaktsioonile vahendaja. Üks katalüsaaa
1. Palmitiinhappe oksüdatsiooni Hº mõõdetuna kalorimeetris on 9958 kJ/mol. Milline võiks olla sama reaktsiooni Hº elusrakus: a) sama Pikemalt: Entalpia on olekufunktsioon ehk sõltub ainult süsteemi olekust, mitte selle saavutamise viisist. (Kips on kips! ja 5=100:5 15=1041) 2. Vette asetatud jäätükk sulab. Miks ei ole võimalik olukord, kus jäätükk muutuks veelgi külmemaks ümbritsev vesi aga soojemaks? Sest isevooluliselt liigub soojus alati soojemalt kehalt külmemale (termodünaamika II säädus) 3. Vee jäätumisel tema korrapära kasvab (S < 0). Kuidas on võimalik vee jäätumine? Vee jäätumisel tema korrapära kasvab ehk S<0. Avatud süsteemi isevoolulised protsessid toimuvad vabaenergia vähenemise suunas (G<0). Selleks,et G oleks negatiivne, peab H<0 ning seega tingimuseks on see,et protsess peab toimuma madalamatel temperatuuridel H>TS Entroopia vähenemist peab kompenseerima soojusvahetus ümbritseva keskkonnaga ja seega peab ümbritsev keskkond
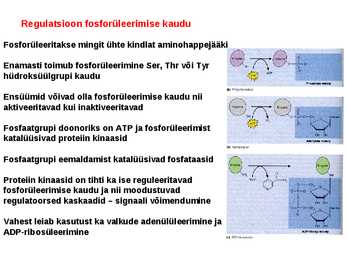
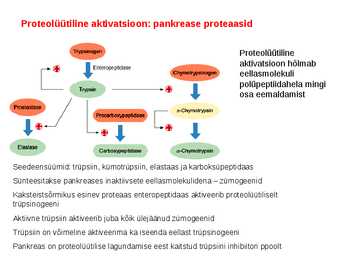
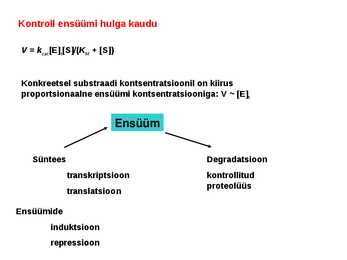
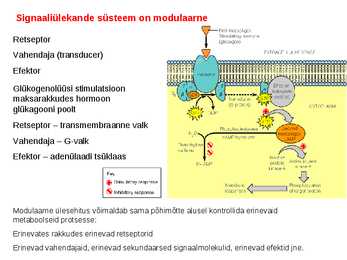
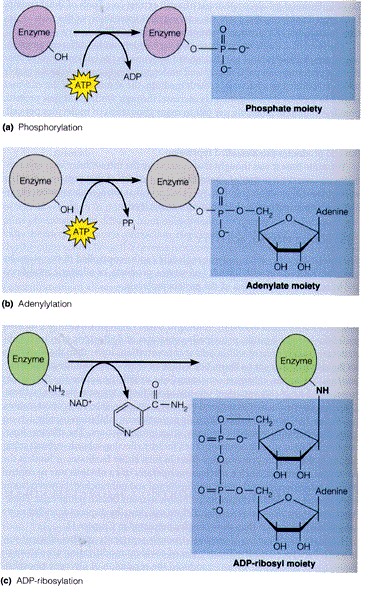
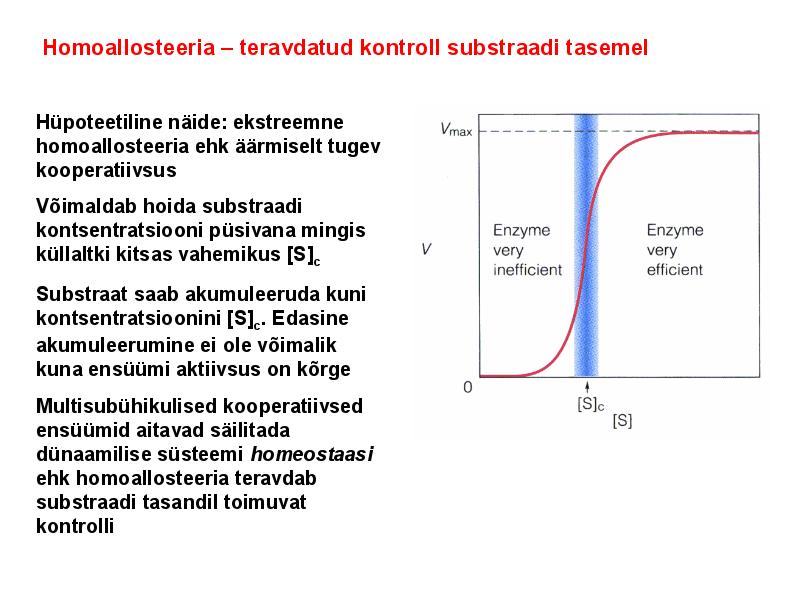
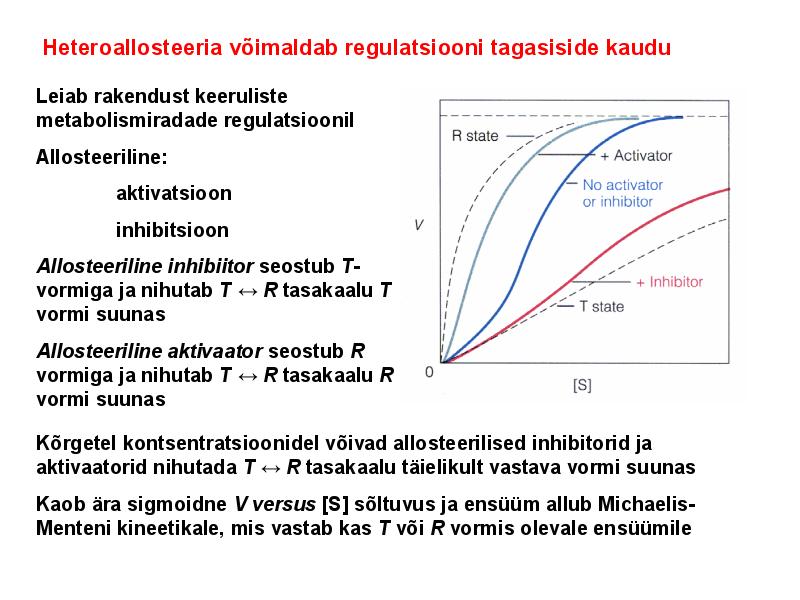
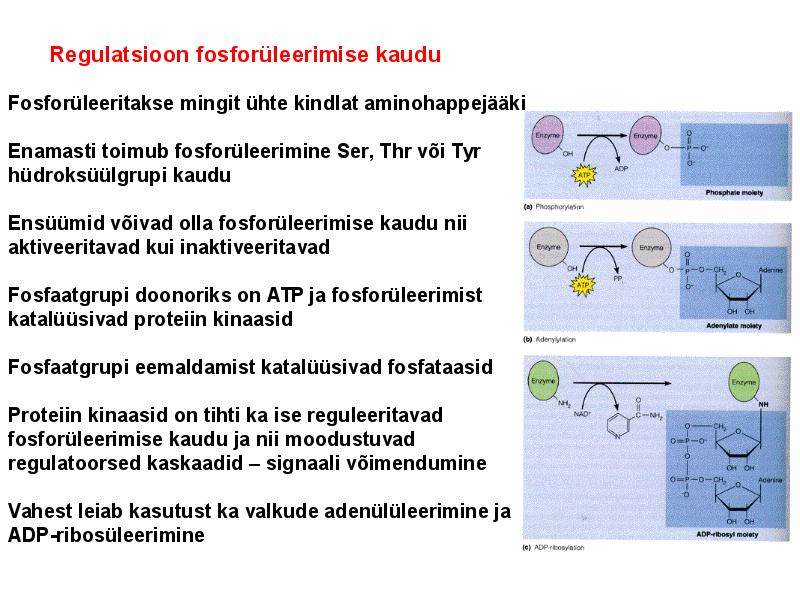
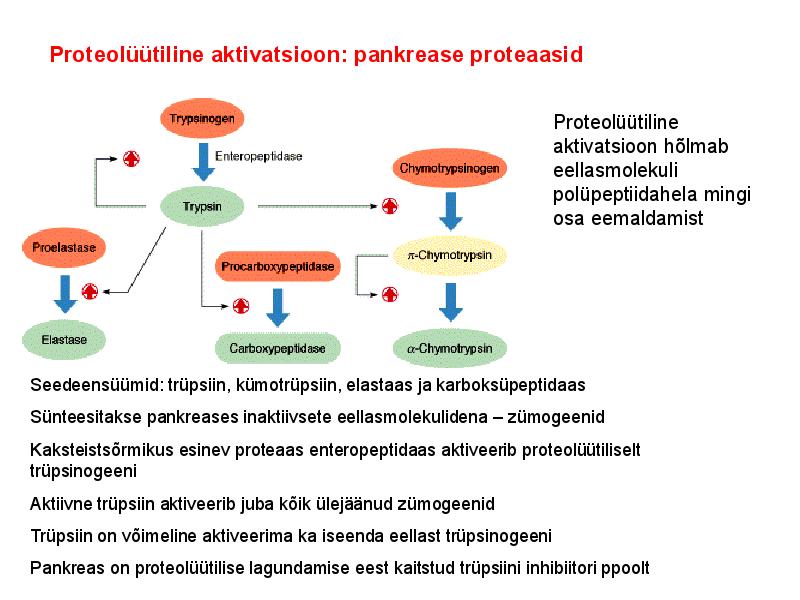
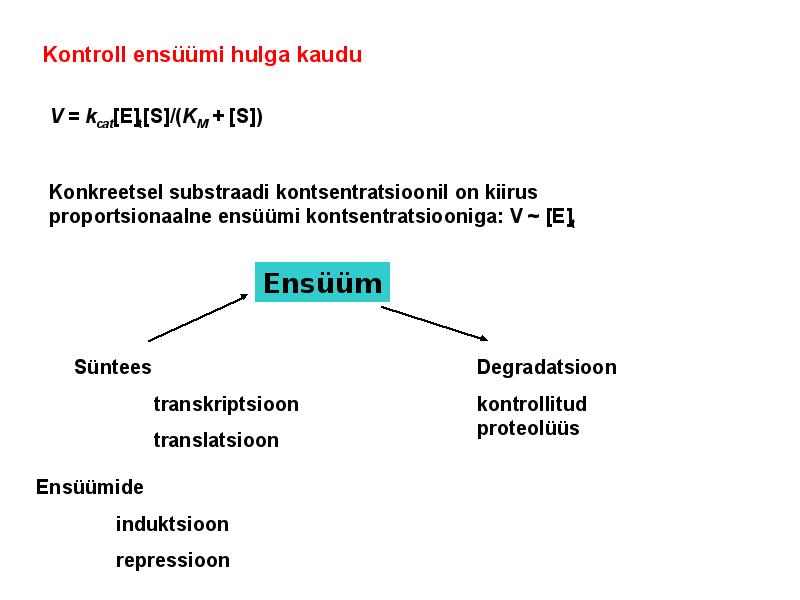
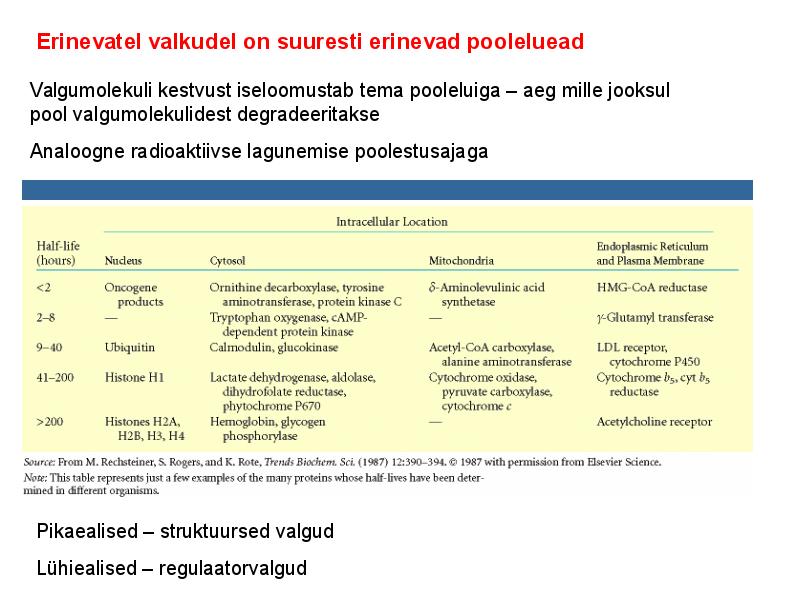
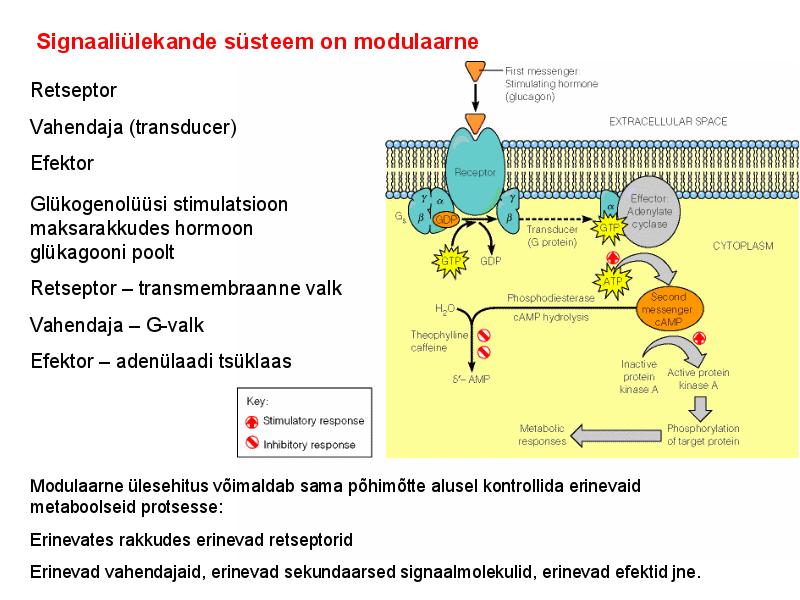
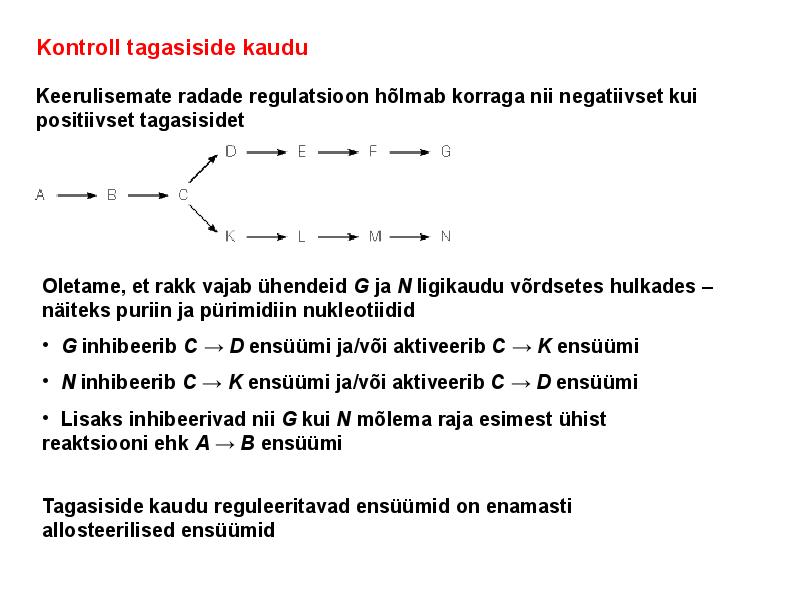
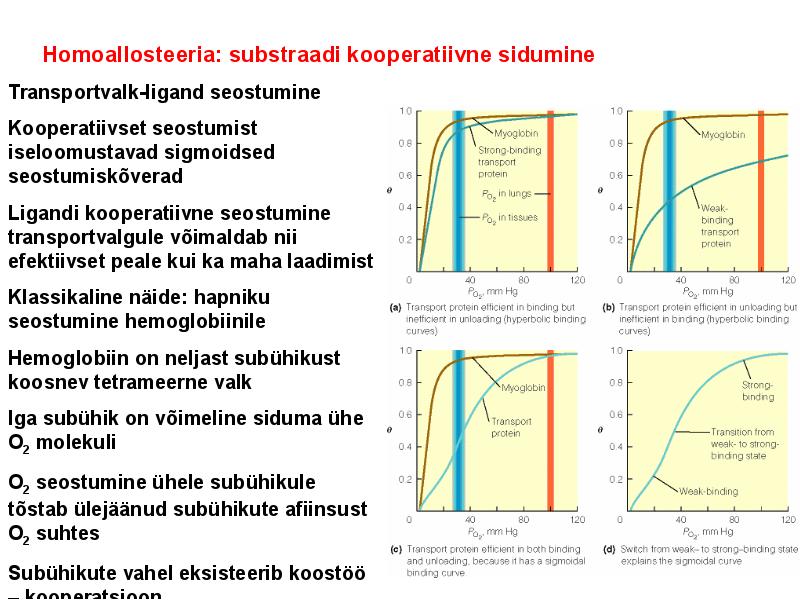
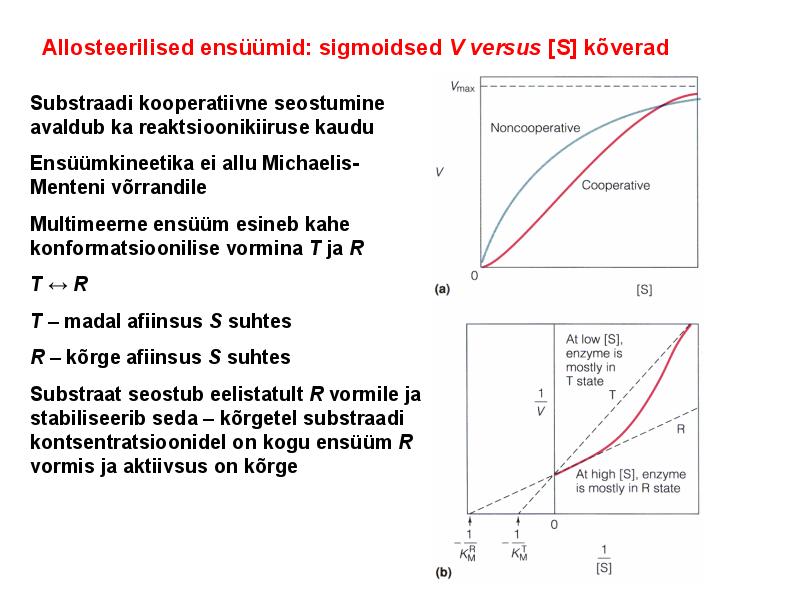
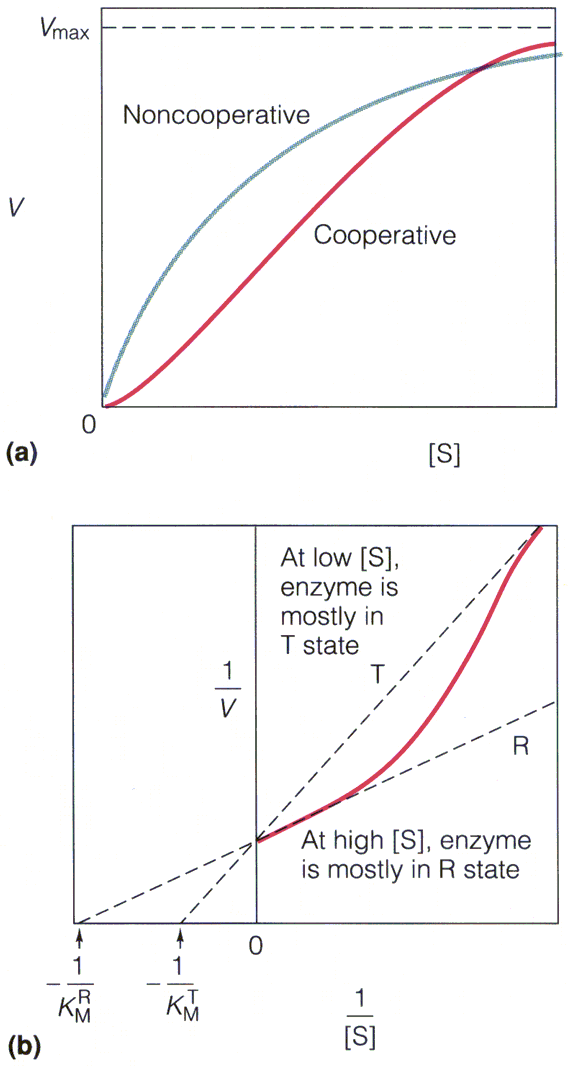
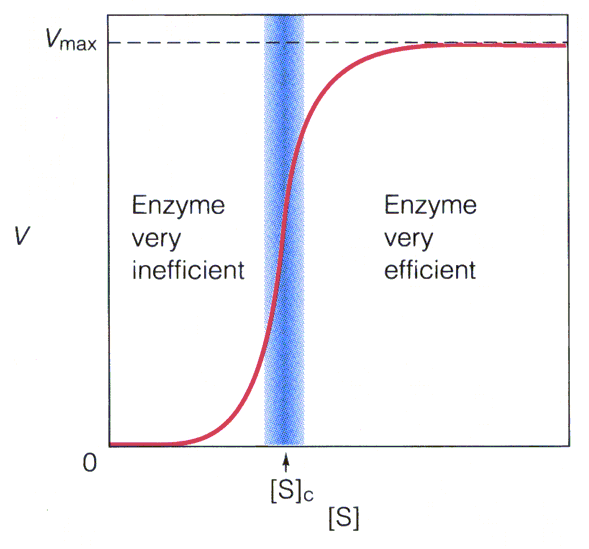
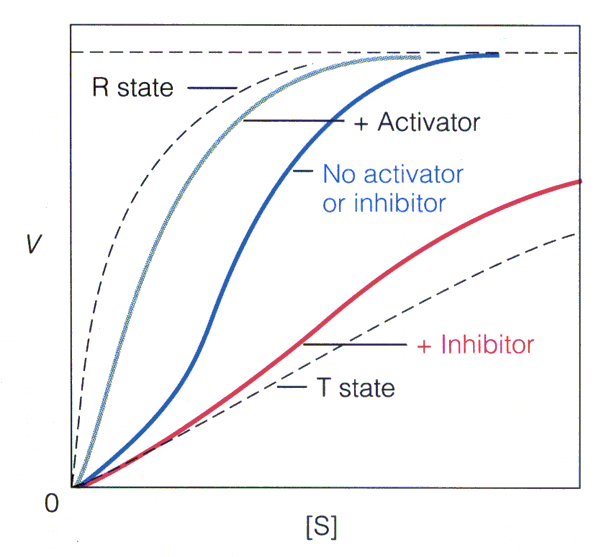
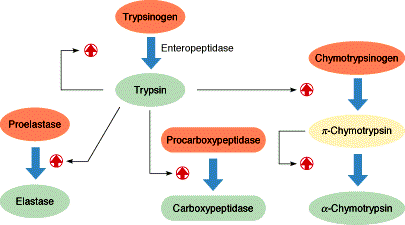
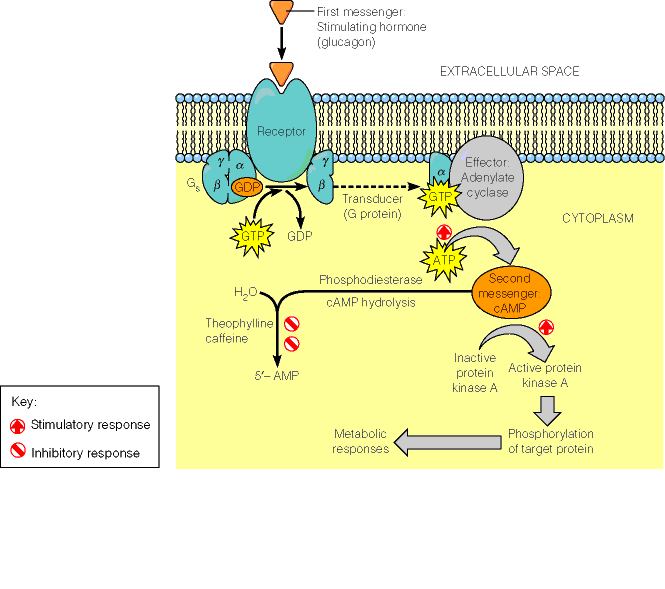
. KATALÜÜSI REGULATSIOON 1) Ensüümide spetsiifilisus milles avaldub ja millele baseerub. Aktiivtsentri mõiste molekulaarne sisu. Stereo-, geomeetrilise, absoluutse spetsiifilisuse iseloomustus. Ensüümi spetsiifilisus on ensüümidele omane võime eristada substraate, millele nad toimet avaldavad. Ensüümide spetsiifilisus toimub molekulaarse äratundmise kaudu, mille aluseks on ensüümi aktiivtsentri ja substraadi struktuurne komplementaarsus. Aktiivtsenter ensüümi molekuli piirkond, mis otseselt osaleb katalüütilises protsessis. Seal paiknevad aminihappejääkide katalüütilised rühmad, mis seovad endaga substraadi. Stereospetsiifilisus võime toimida vaid teatavale stereoisomeerile. Geomeetriline spetsiifilisus võime eristada supstraate molekulis. Absoluutne spetsiifilisus toime avaldub vaid ühele substraadile. 2) Reaktsioonikiiruse reguleerimise võimalused rakkudes. Ensüümide kovalentne modifitseerimine. Valkude fosforüleerimise/defosforül
Ensüümkineetika Michaelis-Menteni võrrand Minimaalne ehk lihtsaim ensüümkatalüüsitava reaktsiooni skeem on kaheastmeline reaktsiooniskeem: 1. Seostumine ensüümi E ja substraadi S vahelise kompleksi ehk ensüüm substraat kompleksi ES moodustumine (pöörduv) 2. Keemiline etapp produkti P moodustumine ensüüm substraat kompleksist (pöördumatu) ja ensüümi vabanemine Produkti moodustumise kiirus V on antud seosega: V = d[P]/dt = k2[ES] Küsimus kuidas sõltub produkti moodustumise kiirus substraadi kontsentratsioonist [S] antud ensüümi kontsentratsioonil [E]t ? Michaelis-Menteni võrrandi tuletamine kiire tasakaalu eeldusel, I NB! Otsime ES kompleksi kontsentratsiooni [ES] sõltuvust substraadi kontsentratsioonist [S] Kiire tasakaalu eeldus: ES kompleksi lagunemine vabaks ensüümiks ja substraadiks on palju kiirem kui lagunemine vabaks ensüümiks ja produktiks (k-1 >> k2) Sellisel juhul on reaktsiooni esimene aste tasakaalus ja me saame
PRIMAARSTRUKTUUR. VALKUDE ISELOOMUSTUS JA BIOLOOGILINE ROLL 14 VALKUDE RUUMILISED STRUKTUURID 16 SISSEJUHATUS ENSÜMOLOOGIASSE 21 ENSÜÜMIKINEETIKA 25 ENSÜÜMIKATALÜÜSI KEEMILISED MEHHANISMID 30 KATALÜÜSI REGULATSIOON 34 MOLEKULAARMOOTORID 43 LIISI KINK 2 BIOKEEMIA test I I. BIOKEEMIA AINE. RAKU EHITUS 1. Bioelemendid. Bioloogilised makromolekulid











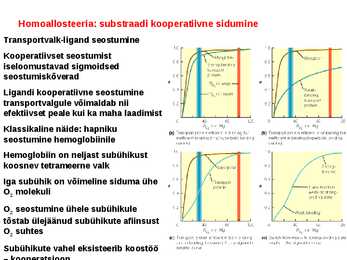
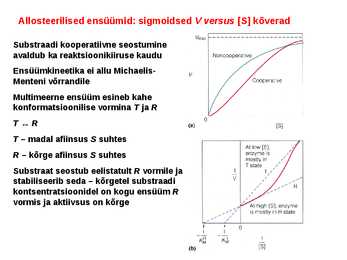
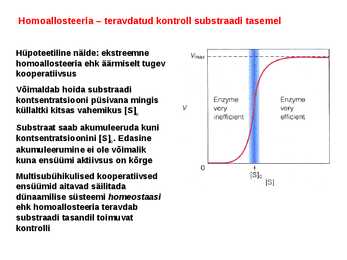
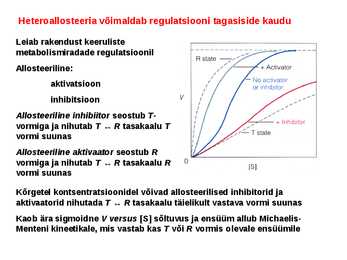
































Kõik kommentaarid