Molekulaarbioloogia võeti kasutusele, et tähistada molekulide struktuuride uurimist
(bioloogiliselt oluliste makromolekulide uurimine), 50ndatest aastatest alates
Objektid, mida käsitleme on samad, mis biokeemias ja geneetikas
Geneetiline teave e pärilik info säilitatakse DNAs nukleotiidse järjestuse kujul. Pärilik teave
säilitatakse nukleiinhapetes nukleotiidse järjestusena.
Valkude liikide nii kvalitatiivne kui ka kvantitatiivne kogu (kuju?) määrab ära organismi
fenotüübi.
RNA
RNA ja RNAga toimuvad sündmused on oluline protsesside kogum. RNA jaguneb kaheks liigiks:
stabiilsed RNA liigid ja
labiilsed RNA liigid. Stabiilse hulka kuuluvad eelkõige pre(rRNA, tRNA,
snRNA), võivad olla väga lühikese elueaga. siRNA (osalevad teiste RNAde metabolismis) ja
miRNA (avastatakse juurde pidevalt). Labiilsed on eelkõige mRNA ja tema derivaadid.
Protsessimise etapp, mis hõlmab järjestuse muutmist, splaissimist, nukleotiidide
modifitseerimist jne. RNA protsessimise juures mängib väga suurt rolli snRNA (väikesed tuuma
RNAd), mis suunavad kohti, kus kohas pre-mRNAga muutused toimuvad (nt splaissing) ja teised
stabiilse RNA liigid osalevad mRNA translatsioonil.
Molekulaarbioloogia dimensioon
: keskne termin on ongström = 10-10 meetrit. Molekulid
ühendatud sidemega, mis jäävad 1-2 ongströmi vahele. Tuumapoorid on u 100 nm e 1000
ongströmi.
Molekulides on aatomid omavahel seotud kovalentsete sidemetega, mis on aatomite vahel olev
tugev interaktsioon. Tugevad interaktsioonid on need, mis hoiavad aatomeid üksteise küljes ja
mis moodustavad molekulide primaarstruktuuri. Molekulide ruumilise struktuuri määravad
nõrgad sidemed: vesiniksidemed, elektrostaatilised interaktsioonid, van der Waalsi jõud,
hüdrofoobsed interaktsioonid. Hüdrofoobsed on need, mis tekivad keskkonna toimel;
eluprotsessid kõik toimuvad veekeskkonnas; vesi surub endast välja need molekulid, mis vees
hästi ei lahustu, sunnib neid kokku kleepuma.
Molekulide struktuuri määravad tegurid
. Ruumiline struktuur (valk ja RNA) on reeglina
määratud nende komponentide primaarstruktuuriga. Kui ühe molekuli struktuur tekib sünteesi
käigus, siis selle juures tuleb arvestada, et kõik makromolekulid sünteesitakse kindlas suunas
(RNA: 5’-3’ suunas, valk: N-terminus-C-terminus suunas). Ruumilise tekke puhul osalevad
paljud lisafaktorid (kõige olulisemad on molekulaarsed chaperonid e molekulaarsed
lapsehoidjad; RNA puhul RNA helikaasid, mis seondudes RNAga muudab RNA struktuuri). Iga
molekuli lõplik struktuur on määratud selle keskkonnaga, kus ta lokaliseerub. Makromolekulid
eksisteerivad kompleksides.
Pärilikku informatsiooni, see mis on määratud DNA nukleotiidse järjestusega, sisaldavad
molekulid koosnevad lihtsast selgroost (kõikidel lülidel ühesugune) ja piiratud arvust
külgahelatest (seotud selgroo külge). Valkude puhul selgrooks on peptiidsidemete ahel. α-
süsinik, mille küljes on aminogrupp ja karboksüülgrupp. α-süsinike küljes AHde külgahelad.
Need AHd, mis sünteesitakse ribosoomis, neid on 20+2 (lisa AHd). 20 esinevad praktiliselt
kõikides valkudes, 2 lisa AHd ei esine mitte kõigis organismides (pürrolüsiin ja selenotsüsteiin).
Nukleiinhapped koosnevad pentoos-fosfaat selgroost ja pentoosidega seotud lämmastikalustest
(iga pentoosi esimese süsiniku juures on lämmastikalus).
1
Nukleiinhapete struktuur
60 a tagasi ilmus artikkel, milles kirjeldati kaheahelalise DNA struktuuriahelat – Crick ja Watson
(25. aprill 1953).
Nukleiinhapped koosnevad nukleotiididest. Iga nukleotiid koosneb omakorda suhkrust, mis
tavaliste nukleiinhapete puhul on pentoos. On ka lämmastikalus ja P-hape, juhul kui P-hapet
pole, siis on see nukleosiid. Nukleotiid on hape ja nukleosiid on neutraalne. Nukleotiidid
esinevad looduses võrdlemisi laialt, ei ole mitte ainult nukleiinhapete sees. Esineb väga
paljudes erinevates vormides ja erinevate funktsioonidega, nt kofaktorid (aitavad mitmeid
reaktsioone katalüüsida või aitavad midagi muud ülekanda), NAD ja FAD+ATP, GTP, UTP jne,
signaalmolekulid (mono-, di-, trinukleotiidid, nt tsüklilised on cAMP ja cGMP) ja ka nukleotiidsed
antibiootikumid. Riboosi nimi on anagramm arabinoosist. Desoksüriboos on ilma hapnikuta
riboos.
Nukleotiidid kui kofaktorid. Nikotiin amiid adenosiin dinukleotiid e NAD ja nikotiin amiid
adenosiin dinukleotiid fosfaat e NADP.
Nukleotiidid kui antibiootikumid. Puromütsiin (valgu sünteesi inhibiitor, aminoatsüül 3’-
otsa arabi..; lämmastikalus adeniin, mille küljes AH metoksütürosiin) ja kordütsepiin e 3’-
desoksüadenosiin (DNA sünteesi inhibiitor; sarnane DNA nukleotiididele, vahe selles et 3nda
süsiniku juures ei ole hapnikku).
Fosforhape ja fosfaatestrid. Fosforhape + alkohol (suhkrud, millel on OH-rühm küljes, OH ei
dissotseeru, vaid H) ˃ fosfaat monoester (üks lämmastikhape + üks suhkur). Fosfaat diester – 2
suhkrut + 2 lämmastikalust + 1 fosfaat. Suhkru aatomeid tähistatakse primmiga (’) ja
lämmastikaluste aatomeid C1. Nt C1 ja C1’.
Nukleiinhappe süntees on polükondensatsiooni reaktsioon, kus eraldub vesi.
Fosfaadid on nukleosiidide küljes erineval moel, nt 3’-monofosfaat, 2’-monofosfaat, 5’-
monofosfaat, aga 1’-monofosfaat ei saa olla (kuna seal küljes peab olema lämmastikalus). Kui
on ühe suhkruga seotud rohkem kui 3 fosfaati, siis on need fosfaadid nt 2’ ja 3’ küljes.
Suhkur
. Suhkru konformatsioon määrab ära selle, missugune nukleiinhappe ahel välja näeb.
Pentoos koosneb 5st süsinikust. Suhkrutele on omane see, et nad esinevad kahes vormis:
lineaarne vorm ja tsükliline vorm. Pentoosid moodustavad 5-lülilise tsükli, mis saab
moodustuda kahte moodi (D või L isomeer – kummale poole jääb OH C-C teljest). Tsüklis sees
olev hapnik on 4nda süsiniku hapnik. Riboosi C2’ hapnik tahab reageerida fosfaadiga ning
selleks piisab mõne raskemetalli lähedusest.
DNA on keemiliselt stabiilne ja RNA on labiilne, mis laguneb kergesti (kerge ka ensümaatiliselt
lagundada). Vahendaja ei tohi olla iseseisev ja seega ka stabiilne. Neil peab olema võimalik
vahet teha (riboos ja desoksüriboos).
Suhkruid võimalik keerata tsükliks kahel erineval moel: kas vasakult või paremalt poolt.
Tekivad peegelisomeerid (D-isomeer kui OH-d on paremal pool ahelat ja L-isomeeril kõik
vasakul). D-isomeer on looduslik isomeer. D-isomeer kui lämmastikalus jääb paremale ja L-
isomeer kui vasakul.
2
H-side. Nukleiinhapetes on erakordse tähtsusega vesinikside. H-sideme puhul on üks H-doonor,
see mis osaleb H-sideme moodustumises loovutab oma elektroni vesinikule. H-sideme eluiga
väga lühike, mis ei kesta kauem kui 10-11 sekundit. H-sideme moodustumisel on alati doonor ja
aktseptor. Doonor – see molekul või aatom, mis annab vesiniku, et tekitada H-side. Aktseptor –
aatom, millel on vabaorbitaal, nt hapnik. Moodustub prootoni ja vabaorbitaali ühine orbitaal.
Doonorite ja aktseptorite paarid on ruumiliselt komplementaarsed.
Watson-Cricki aluspaarid
. Need paarid, mis esinevad tavaliselt DNAs. Amino vormis olev
adeniin paardub keto vormis oleva tümiiniga ja tekib A-T paar, millel on kaks vesiniksidet, mille
vesiniksidemete pikkused on 3 ongströmi. Samasuguse ruumilise geomeetriaga aluspaar tekib
ka guaniini ja tsütosiini vahel, mille geomeetriline sarnasus väljendub selles, et 1’ süsinike
vaheline kaugus on täpselt samasugune; sümmeetriline aluspaar. G-C paar seotud omavahel
kolme vesiniksidemega, võtab enda alla natuke rohkem ruumi.
Watson-Cricki aluspaarid erinevad teistest selle poolest, et nad on sümmeetrilised. W-C puhul
on DNA heeliks alati ühetaoline. Aluspaarid on omavahel seotud H-sidemetega. Kahe fosfaat
selgroo vahele jäävad lämmastikalused. DNA kaksikheeliks kogu aeg pöörab, DNA ahel on
parempidise vindiga. Sealt kust üks ahel läheb teisest üle, sealt keerab paremale. G-C
aluspaarid asuvad enam-vähem heeliksi keskel. Väikeses vaos asuvad osalised aluspaarid ja
osalised desoksüriboosid – seega vähem ruumi. Kahe aluspaarivaheline kaugus on keskmiselt
3,4 ongströmi ja üks täispööre 36 ongströmi (ehk 10,5 aluspaari B-vormis DNA puhul). DNA
diameeter on 20 ongströmi.
A, C, G, T paardumisel on 28 erinevat võimalust. RNA struktuur on ebaregulaarsem,
moodustab erinevaid struktuurielemente, moodustuvad wobble paarid (moodustuvad G ja U
vahel). Wobble paarid on stabiilsed: RNA moodustab rohkem H-vesiniksidemeid (2’ O-d
moodustavad H-sidemeid, mida DNA ei tee + aluspaaride vahelised H-sidemed).
Hoogsteen
paar: moodustub N 6 ja N 7 vahel H-side, moodustuvad teistsuguse geomeetriaga paarid; kuid
põhimõtteliselt on võimalik DNAs ka Hoogsteen paaride põhimõte. Hoogsteen paarid võivad olla
ka ühesugused A-T ja G-C, kuid seal osalevad osaliselt teised aatomid võrreldes Watson-
Crickiga (W-C).
DNA/RNA suur ja väike vagu
DNA kaksikahelale on omased suur vagu ja väike vagu. RNA puhul on ühte vagu (väike)
peaaegu polegi, aga suur vagu on sügav kuid kitsas (ei mahu midagi väga sinna). See, kas 2’
küljes on hapnik, viib suurtele struktuuri erinevustele ja bioloogilistele erinevustele. 2’ hapnik
muudab RNA ahela stabiilsust ja kaksikahela struktuuri.
Geeni regulatsioon põhineb valk ja DNA ära tundmisel – valk ja nukleiinhappe spetsiifiline
interaktsioon. See, mis muudab ahela eriliseks asub suures vaos.
RNA puhul valgud ei saa suurde vakku ligi ning ei saa seostuda. Seostumine ei ole kuigi lihtne,
aga DNA puhul on valkude järjestuse ära tundmine väga lihtne.
RNA A-vormis ja DNA B-vormis, need erinevused põhinevad suhkru erinevatel
konformatsioonidel. Riboos ja desoksüriboos on erinevas konformatsioonis, mis tuleb sellest, et
RNA puhul 2’ süsiniku küljes olev hapnik moodustab vesiniksideme järgmise riboosi 4’
hapnikuga ja see ahelasisene H-side (tekib kõikide järjestikuste ribooside vahel) hoiab RNA
3
kaksikahelat sellise … (aluspaarid teineteisele veidi lähemal, diameeter veidi suurem, puudub
selgelt eristatav väike vagu, ainult suur vagu mis pole eriti kättesaadav)
DNA kaksikahel on painduv, seda saab 10 aluspaari kohta paigutada 18 kraadi ilma lisajõudu
kasutamata. RNA kaksikahel on sirge ja selleks et painutada on vaja aluspaaride vahelisi
interaktsioone laiali lammutada. Kui meie pärilik materjal poleks DNA vaid RNA, siis peaksid
olema rakutuumad e kromosoomid kümneid kordi suuremad, kuna RNAd pole võimalik nii
tihedalt kokku pakkida nagu DNAd. RNAd saab tihedamalt kokku pakkida, aga siis ta ei ole
enam kaheahelaline.
tRNA struktuur on väga stabiilne. RNA struktuure stabiliseerivad lisaks tavalistele W-C paaridel
(kanoonilised tavalised W-C aluspaarid) Hoogsteen ja Wobble aluspaarid ning kolme
lämmastikaluse üheahelaline interaktsioon. Esineb palju rohkem aluspaare ja lämmastikaluste
vahelisi interaktsioone kui seda DNAs. RNA kaksikheeliks vaheldub mitmesuguste struktuursete
elementidega.
DNA kaksikheeliks on enamasti igalt pool ühesugune. DNA struktuuri kaksikheeliksit
stabiliseerivad H-sidemed lämmastikaluste vahel ja stacking. Kuna need on erinevas suunas,
siis H-sidemeid kutsutakse horisontaalseks interaktsiooniks ja stackingut vertikaalseks
interaktsiooniks.
Stacking – elektrostaatiliste ja hüdrofoobsete interaktsioonide kogumik, mis määrab ära
järjestike lämmastikaluste orientatsiooni üksteise suhtes (piki ahelat toimiv jõud).
Hüdrofoobsed tekivad sellest, et nukleiinhape on vesikeskkonnas. Elektrostaatilised on
peamiselt dipool-dipool vastastoimed, mis tekivad seetõttu, et lämmastikalustes tekib
ebaühtlane laengu jaotus, kuigi nad on
aromaatsed kuid mitte täielikult. Väga erineva
tugevusega, mis sõltub järjestusest ja DNA
nukleotiidsest koostisest. Additiivne. Puriin-
puriin˃puriin-pürimidiin˃pürimidiin-pürimidiin.
H-sidemete puhul sõltub stabiilsus ainult
nukleotiidsest koostisest: mida rohkem G-C
paare, seda stabiilsem. Aluspaardumine on
kooperatiivne.
DNA kaksikheeliks
DNA ahelaid saab kergesti lahutada, see on
DNA funktsioneerimises primaarse tähtsusega
(kuna siis on võimalik infot lugeda ja maha kirjutada)
Kaksikahela tekkimist iseloomustab vabaenergia muut e Gibbsi vabaenergia (entalpia,
entroopia ja temp). Vabaen=entalpia muut-temp muut*entroopia (G=ΔH-Δt*S)H-ΔH-Δt*S)t*S)
Kaksikahelaid saab lahutada kergesti, kui tõsta keskkonna temperatuuri – DNA ahelate
lahtisulatamine. Temp, mille juures on pooled ahelad koos ja pooled ahelad lahti nimetatakse
DNA sulamistemperatuuriks Tm
Kaheahelaline DNA võib esineda paljudes erinevates vormides. Enamasti rakkudes on ta B-
vormis. RNA on peaaeg alati A-vormis. B-vormil on suur vagu + väike vagu. A-vormil väikest
4
vagu pm polegi ja suur vagu sügav. Ahel kulgeb sujuvalt, on homogeensed. Kolmas
bioloogiliselt oluline DNA vorm on Z-vorm: üks ahel keerdub üle teise vasakule. Z nimi tuleb
sellest, et ahel teeb Z-kujulise liikumise e kulgeb nagu Z-täht. Z-DNA on võrdlemisi erinev
vorm.
DNA struktuur on väga vajalik. DNA võib olla ruumiliselt väga paljudes erinevates
struktuursetes vormides. Leiti DNA vorm, kus suhkurfosfaat selgroog keskel ja lämmastikalused
on väljapoole pööratud – P-vorm. Mingit tähtsust pole peale ajaloolise, iroonilise.
DNA topoloogia
DNA on nagu kummipael, mida saab venitada pikemaks ja keerata vinte peale-maha. Rakus on
haploidne DNA 3*109 aluspaari. Igas rakus on haploidne DNA u 1 m ja diploidne 2 m. Igas rakus
on siis 2 m DNAd, mis on sinna ära mahutatud niimoodi et iga üksikut osa on võimalik kätte
saada.
Kui DNA ahelat lahti arutada, siis teise kohta tekib vinte juurde – tekivad topoloogilised pinged.
Vintide arv kokku ei muutu, peab jääma samaks (määratud DNA struktuuriga). DNA on
organiseeritud rakkudes topoloogilistesse domäänidesse. Bakteritel on kindlates vahemikes
DNA kinnitatud membraanile, kahe kinnituspunkti vahele jääb topoloogiline domään. Kõik
topoloogilised pinged tuleb lahendada.
210 bp suurusel DNAl on 20 täispööret, L – ahelate seotus, W – (writhing) teise astme vindid.
Kui vähendame täispöördeid, oleme vähendanud ka ahelate seotust ja nüüd see
kompenseeritakse teise astme vintidega. Kui keerame parempidi pöördeid välja, tekib juurde 2
teise astme vinti, mis pööravad samuti paremale. Kui keerame vinte peale, tekivad juurde
vasakpidised vindid. DNA tahab olla alati oma struktuuriga. Kui ahelaid lahutame, tekib sama
probleem – ühest kohast tõmbame ahelat laiali, teises kohas tekivad topoloogilised pinged
(teise astme vintide juurde keeramine või maha). Kui meil on B-DNA, mis on ülekeeratud, siis
kui selles DNAs tekitada ühes väikeses osas Z-vorm, saame topoloogilisi pingeid vähendada.
DNA topoloogiliste muutuste juurde käivad ka topoisomeraasid. Ensüümid, mis muudavad DNA
topoloogiat niimoodi, et tekitavad DNA ahelas katkemise ja seejuures DNA ahel saab ümber
teise pöörelda e ahelate seotuse number muutub. DNA topoisomeraas I tekitab DNAs
üheahelalise katkemise, muudab ahelate seotust 1 ühiku kaupa (tehakse fosfodiester side
katki, DNA võtab normaalse kuju tagasi, laseb topoloogilised pinged välja ja seejärel
topoisomeraas ühendab need otsad tagasi, ei vaja lisaenergiat ega kofaktorit; lõdvestab
struktuuri). DNA topoisomeraas II tekitab DNAs kaheahelalise katkemise ja ta muudab DNA
ahelate seotust 2 ühiku kaupa (kui vaja DNA rohkem kokku pressida e tihemini kokkupakkida,
tekib kaheahelaline katke, DNA ahel saab läbi minna ja tekib selline kaksikahel, kus on üks
supervint sees; seda supervinti välja ei saa ilma ahelate katki tegemisest (ühe ahela katki
tegemisest ei aita)).
DNA topoloogia põhivalem
: L=T+W. Ahelate seotus on numbriline suurus, pole vektorit.
Täispöörded ja vindid on alati vektoriaalsed seega neil on suund ja suurus. T – täispööre, W –
supervint, L – ahelate seotus.
DNA topoloogial on oluline seos mitmesuguste bioloogiliste protsessidega, k.a. haigustega. DNA
erinevaid topoloogilised vormid on määratud DNA nukleotiidse järjestusega, mida iseloomustab
järjestuse sümmeetria. Võivad olla nt riststruktuurid, mis tekivad siis kui meil on DNAs tegemist
5
pööratud kordusjärjestusega e palindroomiga. Kolmeahelalist vormi iseloomustab
peegelkordused, mis on erinevad kordusjärjestusest. Otsesed kordused võivad tekitada
omamoodi struktuure. Kui G nukleotiide on järjestuses palju, võivad need tekitada neljaahelalist
DNAd: struktuur, mis esinevad telomeerides (üks DNA ahel paardub iseendaga). 4 G nukleotiidi
on omavahel paardunud – tetrapleks. Z-DNA saab tekkida sellises DNAs, kus on puriin-
pürimidiin vaheldumisi ja seda vähemalt 10 aluspaari. Z-DNAst B-DNAks saab minna üle väga
kergesti, vaja selleks võtta ainult üks aluspaar lahti.
Kromatiini struktuur
Kaheahelaline DNA on kokkupakitud histoonidega nukleosoomidesse. Iga nukleosoom sisaldab
1,65 pööret DNAd, 147 aluspaari. Nukleosoomi läbimõõt on u 11 nm. Ei paikne ruumis vabalt
vaid moodustavad järgmise astme struktuure, mis võivad olla solenoidid või sikk-sakk vindid.
Moodustavad selliseid struktuure, mis on 30 nm läbimõõduga fibrillis, mis moodustavad
järgmise astme vinte, mis koosnevad lingudest (üksteise küljes, ühendatud kromatiini teljega,
mille moodustab valgud ja RNA). 300 nm struktuur näha valgusmikroskoobis, kondenseeritud
kromosoom 700 nm, mis omakorda moodustab kokku mitootilise kromosoomi 1400 nm.
200 aluspaari, et DNA saaks pöörata 360 kraadi (terve ringi), otsad ei saa ikka kokku –
topoloogia. DNA on nukleosoomis kogu aeg pinge all, rohkem keeratud kui peaks.
Nukleosoomis DNA struktuur ei ole ideaalne W-C struktuur.
DNA ahelad on omavahel seotud vesiniksidemetega, nukleotiidid piki ahelat seotud
fosfodiestersidemega.
Nukleosoom
DNA on pakitud nukleosoomidesse koos histoonidega. Kui toimub DNA replikatsioon,
sünteesitakse uus DNA ja tekivad ka uued nukleosoomid. Kuna DNA süntees on
poolkonservatiivne (mõlemad uued DNA ahelad sisaldavad vana+uue DNA molekuli), osad
vanad nukleosoomid jäävad seotuks vana DNAga …
Nukleosoom koosneb histoonide kehast ja kaheahelalisest DNAst, mis on nagu ümber pooli
keritud. Nukleosoomis on 8 polüpeptiidi ja neid 8 polüpeptiidi on nelja erinevat tüüpi. Igat valku
on 2 koopiat ja need on H2A, H2B, H3, H4. Need neli erinevat histooni on organiseeritud
kahekaupa. Histoonid H3 ja H4 moodustavad tetrameeri (kogu aeg koos, 4 polüpeptiidi), H2A ja
H2B moodustavad kaks dimeeri. Oluline nukleosoomi assableerimise ja organiseerimise puhul.
N-terminaalsed ja C-terminaalsed üheahelalised järjestused, mis ulatuvad nukleosoomi kehast
välja. Need üheahelalised osad on seondunud ühelt poolt DNAga ja teiselt poolt seonduvad
kõikide nende faktoritega, mis aktiveerivad geeni aktiivsust. Esimene märklaud, millega geeni
regulaatorvalgud seonduvad.
DNA on nukleosoomis 1,65 pööret ja see on jaotatud 147 aluspaari vahel. Seega DNA on
veidikene üle keeratud. DNA ja histoonid on seotud omavahel põhiliselt H-sidemetega. H3 ja H4
on seotud selle piirkonnaga, mis tuleb nukleosoomi sisse ja mis läheb välja ehk DNA otstega.
H2A ja H2B on seotud „kerega“, väiksem tähtsus nukleosoomi moodustumisel. H3 ja H4
hoiavad DNAd nukleosoomis kinni ja määravad ära nukleosoomi struktuuri.
Nukleosoomil on sümmeetriatasand e peegelsümmeetria. Rotatsiooniline sümmeetria.
6
H1 on seotud nukleosoomide vahelise DNAga, sellega mis ei ole seotud nukleosoomiga.
Nukleosoomide vaheline osa on erineva ulatusega. Nukleosoomid on kromatiini pakitud
järgmise astme vindina. Moodustades nukleosoomi moodustavad 30 nm fiibri ja nukleosoomid
moodustavad spiraali. 30 nm fiibri struktuuri kohta on erinevaid mudelid (solenoid ja sikk-sakk),
kus mõlemad mudelid on õiged. Seal, kus nukleosoomid on tihedamalt DNA peal (nuklesoomi
vahelist ala vähem), seal on solenoid, ja seal, kus nuklesoomid on hõredamalt DNA peal, on
sikk-sakk.
DNA replikatsiooni ajal toimuvad mitmesugused sündmused lisaks DNA sünteesile: DNA
kaksikahel harutatakse lahti, sünteesitakse uus ahel, osalevad erinevad valgud (helikaas,
topoisomeraas, DNA polümeraas, libisev klamber (DNA polümeraasi β-subühik)). Replikatsiooni
masinavärk e replikatsiooni vabrik seisab pigem paigal ja DNAd sikutatakse läbi. Selleks, et
DNAd sünteesida, tuleb nukleosoomid DNAlt eemaldada: replikatsiooni käigus H3 ja H4 N-
terminaalsed sabad (kõige rohkem modifitseeritud) jäävad DNAga seotuks (suudavad
masinavärgi koos oma modifikatsioonidega läbida ja jäävad täpselt samale kohale, kus nad olid
ja hakkavad moodustama uut värsket nukleosoomi), H2A ja H2B lahkuvad. H3 ja H4 saavad
seotuks jääda nii, et osa jääb ühe uue tütarahela külge ja osa vana ahela külge – jäävad
seotuks mõlema uuesti sünteesitud DNAga.
Uue nukleosoomi kokkupanek on keeruline protsess, kuna on vaja tekitada hulk uusi valk
interaktsioone ja uusi DNA-valk interkatsioone (DNA vaja kokku painutada ja valgud sobitada
kokku üksteise suhtes õigesse asendisse). Selleks on histoonide chaperonid e histooni
lapsehoidjad. Valgud, mis aitavad histoonidel moodustada oma nuklesoomis õiget struktuuri.
H3 ja H4 ühel ahelal ei ole vaja chaperone – tetrameer on moodustunud. Uuesti sünteesitud
tütarahelale on vaja panna H3 ja H4 peale. Neid aitab DNA peale panna õiges orientatsioonis
Asf1 – uuesti sünteesitud valgud, millel ei ole ühtegi modifikatsioone. PCNA replikatsiooni
masinavärki kooshoidev kompleks. Ühel molekuli on vanad H3 ja H4 (nende küljes igasuguseid
keemilisi märke küljes) ja teisel uued H3 ja H4.
Histoonid koosnevad α-heeliksitest. DNA on igalt poolt histoonidega seotud.
DNA replikatsioon
DNA sünteesi viib läbi ensüüm DNA-sõltuv DNA polümeraas. Erakordselt kiire, sünteesib DNAd
kiirusega kuni 1000 aluspaari (1000 fosfodiestersidet sekundis) ning erakordselt täpne. DNA
polümeraas teeb 1 vea võib-olla 1 miljoni õigesti sünteesitud aluspaari kohta. Suudab
katalüüsida mitmeid erinevaid reaktsioone – harutab DNA ahela lahti, parandab oma vigu,
sünteesib aluspaare, paneb uuesti kaksikahela kokku jne.
DNA sünteesil on substraadiks ühelt pool DNA ja teiselt poolt desoksünukleosiid trifosfaat
(dNTP) – alfa, beeta, gamma fosfaat. Selleks, et DNA süntees saaks alata, on vaja vaba 3’ otsa.
DNA sünteesi saab alata ainult olemasolevast DNAst. DNAl peab alati olema matriits, mille järgi
saab sünteesida (peab saama kuskilt selle järjestuse, millel peab olema ka praimer). Praimer-
matriits ühendus – 3’ otsast hakatakse DNAd juurde sünteesima. Praimer on tavaliselt RNA,
sest RNAd on võimalik sünteesida ilma praimerita.
Süntees ei ole keeruline. Matriits määrab ära nukleotiidi, mis järgmiseks lülitatakse ja DNA
polümeraas sünteesib uue fosfodiestersideme, kusjuures järgmine nukleotiid lülitatakse
eelmise nukleotiidi 3’ hapniku külge. Beeta ja gamma fosfaat lahkuvad pürofosfaadi kujul.
Reaktsioon muudetakse pöördumatuks tänu pürofosfataasile, mis hoiab pürofosfaadi taseme
7
väga madalal (pürofosfaat lagundatakse fosfaatideks). Substraadi välja valimine toimub
komplementaarsuse alusel. Seda, kuidas see ensüüm on fantastiliselt täpne on oma kindlad
struktuurse alused.
DNA polümeraase on erinevaid tüüpe, mis sarnanevad oma ruumiliselt struktuurilt parema
käega. Aktiivtsenter (see kus DNA süntees toimub) on peopesas. Aktiivtsentris on praimer ja
matriits. Välja valimine toimub geomeetrilise printsiibi alusel, kus valitakse välja selline
nukleosiidtrifosfaat, mis mahub ära aktiivtsentrisse ja mille alfa-fosfaat satub 3’ hapniku
lähedusse – saab sünteesida nende vahele keemilise sideme. Ribonukleosiidtrifosfaatide
kontsentratsioon on rakus kõrgem kui desoksünukleotiididel (10-20 mikromolaarne, ligi 100x
väiksem). Õige substraadi välja valik toimub aluspaari geomeetria alusel. Keemilise katalüüsi
juures osalevad kaks kahevalentset metalliooni, magneesium. Magneesium A osaleb 3’ hapniku
deprotoneerimisel e oksüaniooni tekitamisel (prooton seotakse metalliooni abil DNA
polümeraasiga; oksüanioon on hästi reaktiivne rühm, mis reageerib peaaegu kõigega – tekib
uus fosfodiesterside), magneesium B hoiab beeta ja gamma fosfaate paigal koordinatiivsete
sidemetega. Oksüanioon reageerib alfa-fosfaadiga ja samal ajal sõrmede domäänist pärit
türosiini jääk (aromaatne AH, mille küljes on benseeni tuum + OH-rühm) hoiab stacking
interaktsiooni kleepimisega substraati paigal, et see ei saaks ära pöörata vms. Lüsiini, arginiini
ja negatiivsete fosfaadi hapnike vahel hoitakse beetat ja gammat paigal. Kui fosfodiseterside
on sünteesitud, lasevad sõrmed lahti ja aktiivtsenter vabaneb. Aktiivtsentris on DNA
organiseeritud niimoodi, et matriits-DNA teeb 90 kraadise pöörde. Selle abil satub
aktiivtsentrisse korraga ainult üks nukleotiid. Sõrmed kokku – fosfodiesterside sünteesitakse –
sõrmed lahti – DNA liigub edasi ühe aluspaari võrra.
Teine liikuv element – pöial – hoiab DNAd paigal sünteesitud DNA poolt ja kontrollib sünteesitud
DNA geomeetriat. Kui DNA aluspaar ei vasta W-C geomeetriale siis DNA väikeses vaos ei ole
suhkrud ja fosfaadid samas asendis. Kui on tekkinud mingi muu paar, kajastub see kohe
väikese vao geomeetrias (kui suhkrute vaheline kaugus muutub). Ei kontrolli ainult viimast
aluspaari vaid viimaseid. Kui peaks juhtuma, et geomeetria ei ole õige (väike vagu ei ole
mõõtmetelt õige), siis laseb pöial väikese vao küljest lahti. Kui pöial laseb DNA lahti, saab DNA
hakata liikuma aktiivtsentris. DNA liigub sealt välja niimoodi, et seal kus DNAd sünteesitakse
pole midagi aga selle läheduses on „uus“ aktiivtsenter, mis lagundab DNAd e
eksonukleaasitsenter. DNA lagundamine on väga aeglane. Selle järel alustatakse uuesti DNA
sünteesi.
Kõik nukleiinhapped sünteesitakse alati 5’-3’ suunas!
rNTP ja dNTP eristamine toimub 2’ hapniku järgi. rNTP 2’ hapnik takistab seostumist
aktiivtsentrisse.
DNA süntees on kõrge protsessiivsusega (näitab seda, kui pikalt protsess käib ühe initsiatsiooni
alusel). Kui DNA polümeraas alustab, jätkab ta oma sünteesi nii kaua kui kogu töö on tehtud.
DNA helikaas, SSBd, topoisomeraasid, primaas, DNA polümeraas, libisev klamber, RNAaas H ja
DNA ligaas – komponendid, mis on vajalikud DNA replikatsiooniks. Lisaks osaleb replikatsioonis
veel rida valke ja ensüüme.
DNA helikaasid on klass ensüüme, mis harutavad lahti DNA kaksikahela. Sageli heksameersed
valgud. Koosnevad 6st identsest subühikust, mis moodustavad rõnga, mis paiknevad ümber
DNA kaksikahela ja liiguvad edasi mööda ahelat (harutab lahti). Võivad olla kahesuunalised –
8
võivad harutada lahti 5’-3’ suunas või 3’-5’ suunas. Helikaasil suur hulk ensüüme, mis ei osale
ainult replikatsioonil vaid ka reparatsioonil ja rekombinantsioonil.
SSB – üksikahelalise DNAga seonduv valk. Selleks, et helikaasi lahutatud ahelad ei läheks
kokku tagasi, osaleb selles SSB. Seonduvad mõlema ahelaga (ei lase ahelatel paarduda).
Aitavad ahelat hoida üheahelalisena, vastasel juhul võiksid need üksikahelad seostuda ka mõne
lähedaloleva muu ahelaga, võõraga.
Kui oleme harutanud ahelad lahti, siis ahelate seotus jääb samaks, kuid täispöörete arv muutub
- tekivad topoloogilised pinged, mille tagajärjel tahab DNA krussi minna. Vintide välja
laskmisega tegeleva
topoisomeraasid. Topoisomeraas – DNA struktuuri muutvad ensüümid
(topoisomeraas I ja topoisomeraas II). Neid pingeid tuleb kogu aeg lahendada. ei lase liialt
spiraliseeruda. Eemaldab superspiralisatsioone
DNA primaas – ensüüm, mis on DNA sõltuv RNA polümeraas. Sünteesib RNA praimereid. DNA
süntees vajab alati matriitsi ja vaba 3’ otsa – selleks primaas. Tekitab väikseid RNA praimereid
kasutades ssDNA matriitsina – tekib vaba 3’ ots, saab hakata sünteesima nukleotiide.
DNA polümeraas – DNA sõltuv DNA polümeraas. Kõige kiirem ja kõige täpsem. Suudab
sünteesida kuni 1000 nukleotiidi sekundis ja lisab olemasolevale 3’ otsale desoksünukleotiide.
Tekib kaheahelaline DNA ahel, millest üks on vanem ahel ja teine uus. Sünteesib alati rangelt
ühes suunas – 5’-3’ suunas. Ühe ahela pealt toimub süntees pidevalt – poolkonservatiivne
replikatsioon. Ahelad sünteesitakse erinevalt. Ühte ahelat saab sünteesida nii kaua kuni
matriitsahelat jätkub. Kui uut ahelat sünteesitakse 5’-3’ suunas, siis vana ahelat loetakse 3’-5’
suunas – antiparalleelselt. Juhtiv ahel. Lohisev ahel (lagging strand) / teine ahel – sünteesitakse
ka 5’-3’ suunas, kuid DNA kaksikahela
lahtiharutamine toimub teises suunas. DNA
polümeraas läheb replikatsiooni kahvlist
eemale. Sünteesitakse fragmentidena –
Okazaki fragmendid (1000- pikk); algab RNA
praimeriga. Äärmiselt kõrge protsessiivsusega.
DNA libisev klamber koosneb kahest identsest
valgumolekulist, mis on omavahel ühendatud
niimoodi, et ühe molekuli peal on teise
molekuli saba. Ühenduskohad koosnevad β-
lehtedest, mis omavahel seonduvad väga
spetsiifiliste AHde varal. Need on hüdrofoobsed
interaktsioonid, mille sees on veel üks ioonne
side. Selline interaktsioon, mis vee keskkonnas
surutakse väga tihedalt kokku.
Selles
hüdrofoobses keskkonnas olev ioonne side, mis
on arginiini ja glutamiinhappe vahel, on väga stabiilne, mis ometi ei ole kovalentne side. Saab
kokku panna ja lahti võtta. Seotud ümber kaksikahela – moodustab ringi ümber sünteesitud
DNA. Ei saa rääkida klambri liikumise suunast, kuna üks ahel kulgeb ühes suunas ja teine
teises. Valk-valk interaktsioonide kaudu seotud DNA polümeraasiga. •
Libisev klamber – Dimeerne valk. Sinine ja roheline monomeer(joonisel). Kumbki monomeer
koosneb tervest hulgast antiparalleelsetest beeta-ahelatest(need on stabiilseimad). Klambris on
9
ka beeta-ahelatele lisaks ka hüdrofoobseid interktsioone – hüdrofoobsed AH-d. Need on seal
omaahel koos sellepärast, et see toimub vesikeskkonnas. Alfa-heeliksid klambri keskel hoiavad
selle sees olevat DNA-d paigal.
RNAaas H – H (sõnast hübriid), lagundab hübriidset molekuli. lagundab RNA, mis on
komplementeerunud DNA-ga Lagundab RNAd DNAst (heterodupleksist). RNA praimerite
eemaldamise tõttu tekivad tühikud, mis täidetakse DNA polümeraasi abil, kuid ta ei ole
võimeline neid ühendama. Tekib DNA, kus iga praimeri kohal on katkemine.
DNA ligaas sünteesib puudu olevaid fosfodiestersidemeid ehk ühendab DNA.
Replikatsiooni masinavärk peab toimuma nii, et mõlemad ahelad saaksid enam-vähem
samaaegselt sünteesitud (et teine ahel ei jääks liiga kauaks ootama). Selleks toimub DNA
replikatsioon DNA replikatsiooni vabrikus, kus on 2 DNA ahela polümeraasi, kus üks on juhtiva
ahela jaoks ja teine mahajääva ahela jaoks. Seotud sellise kompleksiga, mida nimetatakse
PCNAks. Väga tähtis komponent on clamp-loading complex – libiseva klambri DNAle asetamise
kompleks. Trombooni mudel – mahajääv ahel liigub edasi ja tagasi.
Probleem: RNAaas H lõikab 5’ otsast ära RNA praimerid ja jääb seetõttu lühemaks kui 3’ ots.
Juhtiv ahel sünteesitakse lõpuni, kuid mahajääv ahel jääb napiks RNA praimeri osa võrra.
DNA otste replitseerimine eukarüootides
Kui vaatame kromosoomi otste struktuuri, siis eukarüootidel on kromosoomi otstes korduvad
DNA järjestused, mis on suhteliselt lühikesed: TTAGGG. Selliseid kordusjärjestusi sünteesib
telomeraas. Telomeraas on ensüüm, mis koosneb 2st subühikust: valk ja RNA. See valk ja RNA
sisaldab endas telomeeri järjestusele komplementaarset järjestust, kuid sisaldab kordust 1,5
korda ehk ainult AAUCCCAAU. Telomeraas töötab niimoodi, et seondub juhtiva ahela
telomeerse järjestusega (mis asub ahela lõpus) ja sünteesib juurde TTA (pikendab kordust).
Pikendatakse seda otsa, mida saab ehk 3’ otsa. Sedasi pikendatakse kromosoomi otsa palju
pikemaks, kui see enne oli. Praimeri pikendamise tulemusena saab ka sünteesida selle osa, mis
jäi pikendamata. Lõpuks tekib ikka selline struktuur, kus on ühe ahelaline vaba 3’otsaga DNA ja
see vaba 3’ ots (üheahelaline DNA osa) peidetakse ära. Selle peitmisel osalevad telomeeriga
seonduvad valgud, mis ei lase teistel valkudel sinna ligi pääseda. Lisaks moodustab see G-rikas
üheahelaline DNA G-kvartette, iseendaga paardudes moodustab 4-ahelalise DNA, kus ots on
neljaahela sisse ära peidetud (vt 2. loengu slaide, kus oli erinevad DNA järjestused).
DNA replikatsioon raku kontekstis
3R paradigma: replikatsioon, rekombinatsioon ja reparatsioon. Kõik sisaldavad endas DNA
sünteesi, kus sünteesitakse uus DNA. Nendes protsessides osalevad samad ensüümid (mitte
kõik, aga osaliselt kattub). Sarnane valkude kompleks, mis katalüüsivad kõiki 3R protsesse ja
need R-id erinevad omavahel selle poolest, et milline polümeraas mis protsessis toimetab.
E.coli
s on 5 erinevat DNA polümeraasi
, millel on erinevad funktsioonid.
Pol I – reparatsioon, RNA praimeri eemaldamine (osalus replikatsioonil, eemaldab kõige
viimased RNA nukleotiidi).
Pol II – reparatsioon.
Pol III – replikatsioon (koosneb 3st põhilisest nukleotiidist, millega seondub veel 9
valku).
10
Pol IV – reparatsioon, aukude täitmine (erineb teistest oma järjestuse poolest ja ka
aktiivtsentri poolest. Kutsutakse Y-perekonna polümeraasiks. Võimeline parandama
sellist DNAd milleks teised pole võimelised. Võimeline oma aktiivtsentris tekitama
Hoogsten paari – viib paratamatult vigade tekkemiseni, teeb ühe vea ~1000 õige
nukleotiidi kohta – üks mutatsioonide allikas. Hea omadus, et nad teevad palju vigu
seisneb selles et need Y-perekonna polümeraasid on võimeline replitseerima selliseid
nukleotiide, mida W-C ei suuda).
Pol V – aukude täitmine.
Pol I ja Pol II koosnevad ainult ühest polüpeptiidist.
E. Coli’s replikatsioon – algab DnaA(+ATP) seondumisel 9 bp aladele. Nüüd tekib ahelate
lahutamine 13bp kohtade juurest. Saab seostuda uus valk – DnaB, see on helikaas(5’3’). sinna
nüüd lisandub primaas, mis paneb praimeri. Nüüd seondub kummalegi ahelale oma DNA
polümeraas ja klambrid jne.
Kui DnaA vabaneb, siis DnaA lagundatakse. Et DnaA-d enam rakus poleks ja seega ei saagi
kohe uuesti niisama alustada replikatsiooni, on justkui kontrolliks. Ehk DNA-süntees sõltub de
novo valgusünteesist(DnaA sünteesist).
Eukarüootide DNA polümeraasid
Pol α – primaas; koosneb 4st polüpetiidist; sünteesib RNA praimereid
Pol β – BER (aluse eemaldamise) reparatsioon; koosneb 1st polüpetiidist
Pol γ – mitokondriaalse DNA replikatsioon; koosneb 3st polüpetiidist
Pol δ – DNA replikatsioon ja reparatsioon; koosneb 2-3st polüpeptiidist
Pol ε – DNA replikatsioon ja reparatsioon; koosneb 4st polüpeptiidist
φ-π – reparatsioon ja hüpermutatsioon; tekitada mutatsioon sellest kohast kus
vaja
Replikoni mudel
Replikaator – replikatsiooni alguspunkt. Replikon sisaldab replikatsiooni alguspiirkonda, millega
seostub initsiaator. Replikatsiooni alguspunktid on erinevad.
E.coli – üks rõngaskujuline kromosoom, ainult üks alguspunkt. Alguspunkt koosneb 2 tüüpi
kordusjärjestustest. Neid kordusjärjestusi nimetatakse
iteronideks. 13+9 kordusjärjestust – A-
T rikkad.
Imetajate viirus SV 40 – alguspunkt sarnane bakteri omale. Ökonoomsed genoomid, et
replikatsioon oleks võimalikult kiire. Veidi pikemad kordused – vähem, pikemaid kordusi –
rohkem – 65 aluspaari.
Pagaripärm – alguspunkt veelgi lihtsam. Replikatsiooni alguspunkt on selline, kus väga
kordusjärjestusi pole, aga on sarnased järjestus elemendid. 4 järjestust elementi, mis määravad
ära replikatsiooni toimumise.
Initsiaatori funktsioonid replikatsiooni alustamisel
Vajalik selleks, et replikatsioon oleks kontrollitud – et ta algaks üks kord rakutsüklis õigel ajal
õiges kohas. Initsiaator seondub DNAga. Initsiaator valk tunneb ära kindla DNA järjestuse, mis
on unikaalne (esineb ainult replikatsiooni alguspiirkonnas, ainult originis). Selleks, et järjestus
11
oleks unikaalne, selleks peab olema ta piisavalt pikk. Kombinatsioonide arv N, 4n – mitu erinevat
kombinatsiooni on võimalik.
Replikatsiooni mulliga seonduvad helikaasid seonduvad ümber mõlema ahela. DnaA-ATP valk
seondub 9-meerse kordusega, DNA kõverdub, tekib replikatsiooni mull. Seondub 13-
meersetega. DNA kaksikahel sulab lahti, seonduvad DnaB helikaas. DnaC aitab helikaasi peale
panna. DnaC lahkub. DnaB helikaas hoolitseb selle eest, et DNA ahelad oleksid lahus. SSBd ka
seonduvad. DnaA eemaldatakse DNA küljest, lõigatakse pooleks, et ta ei saaks DNAga uuesti
seonduda, sest uut DNAd ei tohi enne replitseerida kui uues replikatsiooni tsüklis. Igas
replikatsiooni tsüklis sünteesitakse DnaA uuesti. Bidirektsionaalne replikatsioon – replikatsioon
toimub kahes suunas. DNA repliktseeritakse täies ulatuses.
DNA – desoksüribonukleiinhape, dnaA – geen, DnaA – valk.
Bakterid on võimelised väga kiiresti jagunema, kiirusega 20 minutit – 20 minutiga on bakteri
rakkude arv kahekordistunud (ideaalsetes tingimustes). Selle aja jooksul ei ole võimalik 4,5
miljonit aluspaari replitseerida, kuna aega ei ole, kuigi DNA polümeraas on väga kiire. Selleks
on bakterites nn
sibulakoore replikatsioon. Toimub väga intensiivne valkude süntees – DnaA
sünteesitakse väga kiiresti juurde. Replikatsioon saab toimuda mitu korda ühe rakutsükli
jooksul.
DNA replikatsiooni alustamine päristuumsetes
Alguspunkt koosneb vähematest elementidest kui bakteritel. Eukarüootides on replikatsiooni
alguspunkt lihtne ja neid on ühe kromosoomi peale hästi palju. Kromosoom algab väga
paljudest kohtadest (kui oleks ainult üks, siis läheks väga kaua aega). Replikatsiooni
initsiatsioon on tihedalt seotud rakutsükliga, algab rangelt ainult üks kord rakutsükli jooksul.
Võib esineda nn sibulakoore replikatsioon (pigem viirustel). Pm on süsteem sama. DNA järjestus
– replikatsiooni origin, mis määrab ära replikatsiooni alguspunkti. Replikatsiooni initsiaator e
orc. Kui orc on DNA alguspunkti ära tundnud, siis toimub helikaaside peale laadimine. Need
helikaasid alguses ei lahuta DNA ahelat, vaid seonduvad DNA ahelaga – kutsutakse Mcm’ideks.
Mcm 1 ei kuulu helikaaside hulka. Kui orc ja Mcm on DNAga seondunud, siis seda kutsutakse
pre-replikatsiooni kompleksiks. Selleks, et pre-replikatsiooni kompleks saaks muutuda
aktiivseks ja saaks replikatsioon toimuda, selleks on vaja kinaase. Nendest kinaasidest kõige
olulisem on CDK. CDK on tsükliin-sõltuv kinnas. CDK fosforüülimise tagajärjel toimub pre-
initsiatsiooni kompleksi aktiveerimine, mis läheb üle replikatsiooni kompleksis. Cdk stimuleerib
ja ka pidrudab replikatsiooni algust. Kui Cdk on kõrge aktiivsusega(fosforüülitud), siis saab
aktiveerida ka pre RC.
Mitu kontrollpunkti: 1) orc seondub originiga; 2) orc kompleksiga seonduvad Mcm-id = kõik
inaktiivne; 3) selleks et saada pre-kompleks aktiivseks on vaja panna külge panna fosforhappe
jääk.
Neid origine on tohutu palju. Pagaripärmis on u 50 000 aluspaari järel üks origin. Mitte kõikide
replikatsioonide originidelt ei hakka replikatsioon pihta. Iga origini peal on orc ja Mcm
kompleks, kuid kõik ei aktiveerita korraga. Kui replikatsioon algab kahesuunas, replitseeritakse
ka 2, 4, 1 origini piirkonnad, siis replikatsiooni käigus lükataks orc ja mcm-id DNA pealt ära. Pre-
replikatsiooni kompleksid inaktiveeritakse (sünteesi käigus kõik inaktiveeritud).
12
Eukarüootide replikatsiooni initsiatsioonil osaleb palju valke. Kuigi origin on lihtsa ehitusega, on
replikatsioon palju keerulisem kui prokarüootidel.
CDK stimuleerib ja pidurdab replikatsiooni algust. Pre-replikatsiooni kompleksi aktiveerimiseks
on vaja CDK kõrgema aktiivsusega komplekse. Alguses CKD aktiivusus madal, saab
moodustada uusi prereplikatsiooni komplekse, kui CDK aktiivsus tõuseb ei saa enam uusi
komplekse moodustada, küll aga saab olemasolevaid aktiveerida. G1 faasis moodustub ainult
uusi prereplikatsiooni komplekse juurde.
Replikatsioon lõpeb ter saidis, replikonid lahutab Topo 2. Replikatsioon lõpeb DNA otsas,
tsirkulaarses DNAs on teistsugune lõpetamine. Tekivad sellised struktuurid, mis on üksteisest
läbipõimunud rõngad – katenaadid. Rõngaskromosoomi ühel pool on replikatsiooni origin, selle
vastas on replikatsiooni lõpetamise piirkond. See on ahela spetsiifiline, juhtival ahelal
lõpetatakse ära ja teisel lõpeb iseenesest. Selle koha peal replikatsiooni kahvel lagundatakse,
katenaadid tuleb lahutada. Seda on võimeline tegema ainult Topo2 (topoisomeraas II). Tekib
katke ühte ahelasse ja tõstab selle teisest üle.
Kromosoomi otste replikatsioon: inimesel
o Teise ahela lõpp teistsugune.
o Sinna jääb replitseerimata ssDNA osa, mida ei saa kuidagi
niisama täis sünteesida. Ja iga replikatsioonitsükli jääbki otsast
umbes 15bp tühimik, lüheneb sealt.
o Telomeer – TG-rikas järjestus eukarüootse kromosoomi otsas.
Neid kordusjärjestusi sünteesitakse telomeraasi abil.
o Telomeraas on ensüüm, mis koosneb kahest komponendist – valk
ja rna. Tema rna osa sisaldab sellist järjestust uaacccuaa, mis on
poolteist vastavat kordust telomeeri otsas.
o Telomeeri dna 3’ otsa sünteesitakse telomeraasi RNA järjestuse
alusel see vastav DNA. Just juhtiva ahela otsa.
o Telomeerides olev DNA sünteesitaksegi RNA alusel ja teine ahel
sünteesitakse komplementaarselt telomeraasi poolt sünteesitud
ahelale juhtival.
o Telomeeridel ka aktiivsus, mis vananedes neil väheneb aina.
Kasvajarakkudel pole sellest juttu.
DNA mutatsioonid ja nende parandamine
Punktmutatsioonid – ühe nukleotiidi asendumine. Transitsioonid ja transversioonid. Tekivad
tüüpiliselt replikatsiooni vigadena. Transitsioonide puhul puriin asendub puriiniga A ja G,
pürimidiin pürimidiiniga C ja T. Aluspaar jääb sama pidi. Efekt väikene. Transversioonide puhul
puriin asdendatakse pürimidiiniga ja vastupidi. Punktmutatsioonid tekitavad AH asenduse
valgus enamasti, geeni funktsiooni see tavaliselt ei muuda. Nonsens-mutatsioonid – kui AH
asemel tekib stopp-koodon, siis lugemisraam katkeb, valku ei saa edasi translatseerida, mõju
suurem, tekib vigane, lühike valk.
•
Tekivad eelkõige replikatsiooni (DNA sünteesi) vigadena. Replikatiivsete DNA
polümeraaside täpsus on väga kõrge: vigade sagedus 1/105-1/106
•
Transitsioonid A Û G; C Û T Aluspaar jääb “sama pidi” (Pu-Py®Pu-Py)
13
•
Transversioonid: A Û T; G Û T; C Û G; C Û A Aluspaar “pööratakse ümber” (Pu-
Py®Py-Pu)
•
Tekitavad enamasti aminohappe asenduse
•
Nende mõju geeni funktsioonile on sageli väike või puudub hoopis
•
Suur mõju on nn. nonsens-mutatsioonidel, mis seisneb stop koodoni tekkimises
valgu avatud lugemisraami (ORF)
Deletsioonid - DNA järjestuse ära kadumine, ühe või enama aluspaari kadumine DNAst. Võib
kaduda üks aluspaar või ka väga suur lõik kromosoomist. Palju suurem mõju. Võivad tekkida nii
replikatsiooni vigadena kui rekombinatsiooniga (eelkõige). Viib tavaliselt geeni funktsiooni
kadumisele. Kui kaob ainult üks aluspaar, siis valgusünteesi lugemisraam läheb segi ja ei saa
enam funktsionaalset valku sünteesida(Ka ühe nukleotiidi puudumine viib lugemisraami
nihkumisele ja kogu järgneva valgu järjestuse muutusele). Bakterites võib deletsioon kaasa
tuua kogu operoni välja lülitamise. Võib rikkuda regulatoorse elemendi funktsiooni, nt võib
kaduda replikatsiooni alguspunkt, DNA ei saa enam replitseeruda.
Insertsioonid – DNA järjestuse juurde tekkimine. Tekivad eelkõige rekombinatsiooni tulemuse
või reparatsiooni käigus. Tekivad transposoonide jäljena. Insertsioonid rikuvad lugemisraami
aga samuti regulatoorsete järjestuste funktsiooni.
Mikrosatelliidid – 2-4 aluspaaride kordused, korduvad palju kordi. Nende korduste arv võib
väga kergesti muutuda. Telomeeride ja tsentromeeride alas on neid väga palju.
Kordusjärjestste korduste arvu muutused on sagedasemad insertsioonid ja deletsioonid.
Homopolümeersed piirkonnad on küll haruldasedmad aga insertsioone ning deletsioone on neis
sageli
Peale replikatsiooni värskelt sünteesitud DNA struktuur kontrollitakse mitu korda üle,
kõigepealt DNA polümeraasid väikses vao poolt, vahest ei pane ta viga tähele.
MutS valk, valepaardumise parandamine – homodimeer, mille kaks poolt on omavahel seotud
ümber DNA, mõõdab DNA kaksikheeliksi läbimõõtu, libiseb üle DNA kaksikahela. Kui leiab koha,
kus ei ole läbimõõt 20A, on tekkinud vale paardumine, siis jääb ta seal seisma ning seondub
DNAga palju tugevamini kui normaalselt, hakkab ATPd hüdrolüüsima ning ta struktuur muutub.
Kutsub kohale MutL ja MutH, mis ilma MutSita DNAga ei seondu, nad seonduvad MutSiga. MutH
valk hüdrolüüsib ühe fosfodiestersideme. MutL määrab, kumba ahelat parandada(või mitte
parandada), see valk oskab vahet teha keemiliselt modifitseeritud ahelaga ja mittemod.
ahelaga, MutL tunneb ära metüleerimismustriga ahela(Replikatsioonil tekib hemimetüleeritud
DNA. Sellest metüleerimise mustrist oleneb, kumb lagundatakse – üks ahel on antud juhul
metüleeritud, teine mitte). MutL ja MutH seonduvad erinevate DNA ahelatega. Sinna kuhu
seondus MutH tehakse ahelasse katke, lagundatakse ahel. MutH seondub selle ahelaga, mis ei
ole metüleeritud ehk värskelt sünteesitud ahelaga.
MutS valk, ahela lagundamine
Ta on nukleaas. Struktuur: Koosneb kahest identsest subühikust, mõlemad subühikud
koosnevad 5 domäänist: isomerisatsiooni domään, mismatch domään (osaleb DNA
valepaardumise ära tundmisel), sidumisdomään, ATPaasi domään.. N-terminal mismatch-
recognition domain. Connector domain. Core domain. Clamp domain; the clamp domain has a
beta-sheet structure. ATPase domain. HTH (helix-turn-helix) domain, which is involved in dimer
contacts.
14
Valesti paardunud G-T paar soodustab DNA kõverdumist. Sellega seondunud MutS valk
painutab veel rohkem DNA kaksikahelat. Väike vagu on tunduvalt suurem kui ta peaks olema.
Mut S valk mõõdab väikese vao poolt fosfaatide vahelist kaugust. DNAga kokku jäädes tõmbub
kokku, muutub kompaktsemaks, siis seonduvad MutH ja MutL.
Põhiline funktsioon on parandada replikatsioonil tekkivaid vigu, parandab just uue ahela peal
vigu. MutL valk seondub metüleeritud ahelaga ehk vanemahalaga, MutH valk aga
metüleerimata ehk tütarahelaga. MutH on nukleaas, katkestab ühe fosfodiestersideme. Edasi
mitmete nukelaaside koostoimel lagundatakse vigane piirkond. MutH seostub sinna poole MutS
valku, kus on lähim metüleeritud koht. 5’-3’ suunas lagundavad DNAd kindlad nukelaasid, 3’-5’
suunas on aga hoopis teised nukleaasid. DNA polümeraas sünteesib lagundatud kohta uue DNA
asemele ja ligaas ühendab ahelad.
Restrikitsioonid-modifikatsioonid
•
Sama DNA järjestuse tunnevad ära nii restriktaas kui metülaas
•
Restriktaas ei lõika metüleeritud DNA’d
•
Restriktaas lagundab metüleerimata DNA (võõra DNA) spetsiifilistest kohtadest
Restriktsioonid tunnevad ära kindlaid DNA järjestusi ja lõikavad mõlemad ahelad katki, võivad
seda teha sama koha peal või erineva koha peal ahelates.
Bakterite rakkudes on sellised restriktsiooni mutatsioonid, mis kaitsevad bakterit võõra DNA
eest. Bakteriofaagi DNA on teistmoodi modifitseeritud kui bakteri peremehe oma, siis see aitab
neid eristada. Metülaas – DNAd metüleeriv ensüüm. Restriktaas – DNAd lõikav ensüüm.
Tavaliselt 4 või 6 nukleotiidi pikk see DNA järjestus mida ära tuntakse. Metülaas metüleeriv
DNA ja restriktaas ei lõika metüleeritud DNAd katki ehk siis peremehe oma. Võõras DNA, millel
pole metüleeritud märki lõigatakse katki ehk bakteriofaagi oma.
Modifikatsiooni-restriktsiooni süsteemid bakterites:
o
Bakterite suurimad vaenlased on bakteriofaagid – oma DNA-ga mikroobid.
o
Bakteritel see süsteem võitlemaks bakteriofaagide vastu.
o
Selleks olemas neil ensüümid – restriktaasid. Nad tunnevad ära kindla DNA
järjestuse(4-6bp) ja ta lõikab DNA selle järjestuse kõrvalt või seest
katki(mõlemad ahelad) ning siis sageli järgneb selle DNA lagundamine.
o
Restriktaasid töötavad koos metülaasidega, mis tunnevad sama järjestuse
ära ning metülaas nüüd metüleerib selle.
o
Nüüd restriktaas jätab lõikamata metüleeritud DNA(enda oma) ja lõikab
metüleeritud(võõra DNA) spetsiifilistest kohtadest.
Iga 250 aluspaari järel tuleb sama 4 nukleotiidine järjestus ette, mida metüleeritakse. Uus
sünteesitud ahel metüleeritakse sama moodi nagu vanem ahel.
Reparatsioon
Kui DNAga toimub mingi kahjustus, siis ta kas parandatakse või rakk sureb ära. Hulkraksetes
organismides suunatakse rakk apoptoosi. Parandamine toimub ka RNA tasemel. Kui RNAd ei
õnnestu parandada, siis see lagundatakse, sest RNAd on palju ja saab DNA pealt alati uusi
juurde sünteesida. Sama käib valkude kohta. DNA – kas reparatsioon või degradatsioon(see viib
raku hukkumiseni). RNA – RNA reparatsioon või degradatsioon(rakk ei hukku). Valgu puhul
sama, mis RNA-ga.
15
DNA reparatsiooni mehhanismid
Enne replikatsiooni: vigase nukleotiidi lagundamine (dut, ung)
Repl ajal: kineetiline veerulugemine, toimetamine
Peale replikatsiooni :
A)
nukleotiidi taastamine (metülaasid, fotoreaktivatsioon)
B)
Uue DNA sünteesi abil toimuv reparatsioon (BER, NER, mismatch R, Y-repar, rekomb
repar)
Algab juba enne replikatsiooni. Need nukleotiidid, mis ei vasta standardile, nt
desoksünukleotiidid on desoksüuridiinid(võib olla ainult ribouridiin rakus) keemiliselt
kahjustatud, need lagundatakse. Lämmastikalus lõigatakse suhkru küljest lahti ja lagundatakse
või saadetakse rakust välja. Replikatsiooni ajal toimub ka parandamine. DNA polümeraasil on
võime DNAd lagundada kohe.
Kõige olulisemad reparatsiooni mehhanismid aga pärast replikatsiooni alles. Kaks erinevat liiki:
otsene reparatsioon ehk nukleotiidi taastamine. Ja
uue DNA sünteesi abil ehk
lagundatakse üks kahjustatud ahel ja sünteesitakse uus selle asemel.
Lämmastikalused on keemiliselt aktiivsed. Neid hoitakse DNAs nii, et nad ei puutuks kokku
välise keskkonnaga, et ei reageeriks, kuid ikka juhtub seda. Nt tsütosiin reageerib veega,
tsütosiini neljanda süsiniku juurde tuleb lämmastiku asemel hapnik, muutub uridiiniks. Selle
suudavad rakus olevad ensüümid ära tunda, see lõigatakse välja DNAst ja lagundatakse ära.
See on üks põhjust miks DNAs ja RNAs on erinevad nukleotiidid. Guaniin võib reageerida ka
veega, aga ka väga paljude muude ühenditega. 5-metüültsütosiini deamineerimise tulemusel
tekib tümidiin, mida ei ole võimalik ära tunda enam.
Pürimidiindimeeride teke
: kui DNA ahelas on järjestikku kaks pürimidiini, siis need võivad UV-
valguse toimel omavahel reageerida ja moodustada tsütobutaaniringi. Kui neid tekib hästi
palju, siis epiteeli rakud ei suuda enam neid kahjustusi parandada, rakud lähevad apoptoosi,
tekib punetus ja sügelus.
Etiidiumbromiidiga värvides lähevad värvi molekulid DNA aluspaaride vahele. Ka
akridiinoranž. See muudab DNA topoloogiat, ta paindub. Takistab DNA normaalset
metabolismi. Inihibeerivad topisomeraasi. On DNAle kahjulikud. Bensbüreenid – tekivad
kuumal ilmal auto rehvidest kiirel sõidul.
Biopolümeeride kahjustused, oks. stress: ioniseeriva kiirguse toimel, biosünteesi vead,
keemilised kahjustused. Olulisim on
oksüdatiivne stress, reageerimine hapniku
radikaalidega(Põhjuseks arvatakse, et elu tekkis ju redutseerivates tingimustes, seega pärilikku
informatsiooni kandvad molekulid pole vastuvõtlikud, harjunud hapniku olemasoluga), ka
metaboliitide tingitud kahjustused. Võib toimuda nii DNA, RNA kui valkudega. Hapnikust tekivad
väga kergest vabad radikaalid.
Kahjustuste likvideerimine, DNA parandamine:
vigased molekulid tuleks lagundada. DNAd ei
saa lagundada. Seda tuleb parandada. Ööpäevas kuni 500 000 DNA kahjustust inimesel kes
võtab päikest ja teeb igast muud nalja. Nii tuuma kui mitokondri DNA kahjustub koguaeg. Kui
DNA parandamise kiirus on sama suur kui kahjustuste tekkimise kiirus, kui on tegemist terve
rakuga. Kui aga kahjustusi tekib rohkem kui neid parandada jõutakse on tegemist haige rakuga,
on koormaks kogu organismidele.| On vajalik homeostaasi säilitamiseks, vigased molekulid ei
16
täida oma ülesandeid ja takistavad ka tervetel molekulidel
toimida. Lihtsaim viis vabanemiseks on vigane molekul
lihtsalt lagundada, see aga ei sobi DNA puhul, küll RNA ja
valkude puhul(DNA lagundamine viis ju raku hukkumiseni).
Otsene molekulide parandamine taastab algse seisu.
Inimesel on 130 geeni, mis tegelevad DNA parandamisega.
Mõnel bakteril on kuni 1/3 genoomist DNA parandamisega
tegelev, nt Deinococcus radiodurans. Paljudel suurtel
viirustelgi on oma DNA parandamise süsteem.
DNA reparatsiooni süsteemid: (tüüp-viga-
ensüümid)
1.
Valepaardumine – replikatsiooni viga – MutS,
MutL, MutH
2.
Fotoreaktivatsioon (otsene parandamine) – Py
dimeerir (alküleerimine) – DNA fotolüaas, AlkB
3.
B E R – N-aluse modif – DNA glükosülaas
4.
Nukelotiidide väljalõikamine N E R – (Py)2,
Aluse MOD – Uvr A, B, C, D, XPA, XPC, XPD
5.
2-ahelaliste katkete parandamine (vabad
kaheahelalised otsad tuleb ära peita) – 2-ahelaline
katkemine – RecA, RecBCD, Ku
6.
Rekombinatsiooniline reparatsioon (DNAd ei ole võimalik nende kõigi teiste
asjadega parandada, mõlemas ahelas on viga või üks läheb katki siis kui parasjagu teist
parandad vms) – massilised kahjustused – UmuC (DNA Pol IV ja Pol V)
Fotoreaktivatsioon
– fotosünteesivatel organismidel on eluliselt tähtis saada ioniseerivat
kiirgust, tänu sellele on vaja oma fotosünteesi aparaati eksponeerida väga hästi valgusele. DNA
saab aga ka paraku pihta, taimedel on
ensüüm DNA fotolüaas, mis parandab kahjustusi.
Fotolüaas seondub DNAga, kasutab parandamiseks nähtavat valgust.
Otsene reparatsioon: Alküleerimisest tulenev kahjustus parandatakse metüültransferaasiga.
Alküülrühm, mis on nukleotiidi küljes võetakse ära metüültransferaasi poolt. Tekib metüleeritud
valk, mille saatus on saada lagundatud. Kallis protsess, terve valk läheb raisku. Alfa-
ketoglutaraadi abil saab ka neid parandada. See võtab ka metüülrühma ära, kasutab
koensüümina 2-oksoglutaraati. Tekib formaldehüüd ja lämmastikalused taastatakse algsel
kujul. Nii DNAs kui RNAs töötab.
G nukleotiidi parandamine, ta on keemiliselt kõige reaktiivsem, oksüdeerimine toimub eriti
lihtsalt. Oksüdeeritud G nukleotiid paardub teistmoodi kui tavaline, tema vastasahelasse
lülitatakse A nukleotiid, G-A paar tekib. Selle lõikab välja
glükosülaas.
NVR reaktsioonid – Uvr teema
UvrAB skaneerivad DNA’d
UvrA lahkumisel UvrB
sulatab DNA ahelad lahti
17
UvrC tekitab katked
Helikaas UvrD eemaldab
kahjustatud DNA
DNA Pol I ja ligaas
teevad DNA korda
Reparatsioon on seotud transkriptsiooniga. Need DNA
piirkonnad, millelt RNAd sünteesitakse on pidevalt
RNA polümeraasi kontrolli all. RNA polümeraas jääb
seisma, kui leiab vea, transkriptsioon peatud ja aitab
reparatsiooni mehhanismidel seonduda paremini DNA
peale. Neid DNA piirkondi, millelt toimuba RNA
süntees kontrollitakse sagedamini, kui teisi DNA
piirkondi. Mittekodeerivas piirkonnas parandatakse
vigu vähem.
RNA reparatsioon
Otsene parandamine (AlkB), dealküleerimine
•
tRNA parandamine (CCA, antikoodon lingu tgaastamine)
•
rRNA parandamine.
•
RNA-ga sagedasem juhtum – ahela katkemine
Valkude parandamine
Valgud funktsioneerivad vaid õiges ruumilises konformatsioonis.
Valkude oksüdatiivsete kahjustuste parandamine – karbonüleerimise
parandamine
Metioniini sulfoksiidide redutseerimine – universaalne protsess kõigis
organismides.
Isoaspartaadi parandamine
Homoloogiline rekombinatsioon:
Rekombinatsioon jaguneb kaheks: homoloogiline ja kohtspetsiifiline.
Homoloogiline – Vajab vähemalt identset järjestust 50 bp, siis saab alata, parem on kui neid
on enam. Põhiolemuselt toimub üht moodi nii eu- kui prokarüootides. Homoloogilised
kromosoomid vahetavad homoloogilisi DNA järjestusiˇ. Eukarüootides toimub crossing-over –
kromosoomi ristsiire. Prokarüootide nimetatakse homeoloogiliseks, sest seal peamiselt genoom
haploidne. Homoloogiline rekombinatsioon on väga tähtis stabiliseerimaks genoomi, et selles ei
toimuks reparatsioonidega suurt muutust.
oTransgeenseid organisme konstrueeritakse ka HR abil. Homoloogilise rekombinatsiooni
reaktsioonid
Esimene on homoloogiliste DNA lõikude(alleelide) joondamine, lähestikku
viimine. See pole väga lihtne, sest kromosoom ~1m. Hulkraksetes on väga
palju kordusjärjestusi, seetõttu ka homoloogilisi järjestusi eri kohtades.
18
Siis toimub 2.ahelaliste katkete tekitamine ühes joondatud DNA molekulis.
DNA lagundatakse, tekitatakse 1-ahelalaine DNA
Ahela vahetus ja Holliday ühenduse teke – üks kahest ahelast lagundatakse ära,
tekib üheahelaline DNA. Üheahelaline tungib kaheahelalise sisse, vahetab välja
ahela, tekib Holliday ühendus
Ühendus nüüd liigub, moodustuvad uued aluspaarid. Kuni tekivad
erinevused(valepaardumine), kuni homoloogia lõppeb.
Rekombinatsiooniproduktide lahutamine. Sellest kuidas produktid lahutatakse
sõltub, kas rekombinatsioon tekib või mitte.
Holliday mudel – lihtsustatud, ei ole päris õige
Splice recomination ehk
liit-rekombinatsioon tekivad ristproduktid ehk crossover products
või portsjonrekombinisatsioon (tegelikult rekombineerunud molekule ei teki).
Kaks DNA piirkonda, mis sisaldavad identseid järjestusi, homoloogilised kromosoomid. Holliday
mudeli kohaselt algab rekombinatsioon sellest, et mõlemas DNA molekulis tekitatakse
üheahelialine katkemine identses kohas. Ahelad lähevad omavahel risti, tekib hübriidne DNA
molekul – see ongi Holliday ühendus. Holliday ühendus liigub, aluspaarid samal ajal sulavad
lahti ja kohe tekivad uuesti, kuni jõuab piirkonnani, kus komplementaarsus ei ole enam täielik.
Tekivad mitu lähestikku asetsevat vale paardumist, üksik ei mõjuta, Holliday ühendus jääb
seisma ning lahutatakse. Ahelaid saab omavahel lahutada kahel erineval viisil, vastavalt sellele
kuidas lahutatakse tekivad erinevaid produktid.
Liit-rekombinatsioon
i produktid tulevad siis, kui lõikame katki need ahelad, mida alguses ei
lõigatud. Kui aga lõikame need, mida ennegi lõikasime, siis tulevad
portsjonrekominatsiooni produktid.
RuvA – rekombinatsiooni valk.
RuvB – helikaas, mis kaksikahela lahti
harutab.
RecBCD rada
, ehk tegelikkus:
Rekombinatsioon algab identse järjestusega
DNA piirkondade joondamisega. Toimub ka
katkemine kahes ahelas, aga need ahelad on
pärit samast molekulist ehk siis üks
kaksikahelaline katkemine. Selle
katkemisega seondub kompleks, mis
koosneb kolmest subühikust: RecB, RecC ja
RecD. Esimene etapp on kaksikahela täielik
lagundamine. RecBCD kompleks suudab
lagundada DNAd mõlemas suunas. DNA
lagundamine toimub nii kaua, kuni jõutakse
hii saidini, neid kutsutakse rekombinatsiooni
tulipunktideks. Selle koha peal muudab
RecBCD kompleks oma omadusi, RecD
lahkub, hakkab lagundama edasi ainult seda
19
ahelat, mis lagundati enne 5’-3’ suunas. Selle üheahelalise DNA ahelaga seonduvad RecA valgu
monomeerid, igas RecA filamendis moodustub kaks DNA seondumise piirkonda. DNAga
seondumine toimub fosfaadi hapnike kaudu, järjestusest sõltumatu seondumine. RecA aitab
üksikahelaisel DNAl tungida kaheahelalise vahele, et moodustud Holliday kompleks. RecA
filament liigub mööda kaksikahelalist DNAd seni kuni leitakse komplementaarne koht.
Võrreldes Holliday rajaga toimub siin suures hulgas nii DNA lagundamine kui süntees, DNA
polümeraas sünteesib asemele selle, mis enne ära lagundati.
Helikaasid (RuvB) kasutavad ATP hüdrolüüsi energiat, et ühendust liigutada.
RuvC – nukleaas, mis katalüüsib Holliday ühenduse lahendamist.
Kui juhtivas ahelas on toimunud katkemine, siis süntees peatub seal, replikatsiooni kahvel
laguneb ära. Sellist kaheahelalist katkemist saab parandada RecBCD kompleksiga. Ta
lagundaks ühe ahela ära ja parandaks kohe ära. Kui DNA ahelas on mingit tüüpi kahjustus,
millest DNA polümeraas ei ole võimeline üle sünteesima, replikatsiooni kahvel jääb seisma
sinna. Juhtiv ahel jääb seisma, teist ahelat sünteesitaks edasi. Moodustub kaheahelaline
struktuur ja RecBCD on võimeline seda parandama.
*kui tuleb mingi DNA rakku, kui selles puudub hii-sait, siis see lagundatakse niisama ära, ei
toimu mingit ristsiiret.
Rekombinatsiooni ensüümid
:
Kõige tähtsam ensüüme on E.colis RecA, eükarüootides ta nimi Rad51. Kui see ensüüm puudub,
puudub bakteril võime parandada vigu homoloogilise rekombinatsiooniga. 2-ahelalise katke
tegemiseks on ka Spo11 valk, sama nimi nii bakterites kui ka eukarüootides. Üheahelalise DNA
tekitab RecBCD kompleks.
RecA valk on võtmeensüüm homoloogilisel rekombinatsioonil. RecA filament tekib samas
suunas kui vaba 3’ ots tekib. RecA filamendis on DNA üksikahel samasuguse konformatsiooniga
nagu ta on kaksikahelalises.
Reaktsioon
Ensüüm E.coli
Ensüüm eukarüoot
ahela vahetus
RecA
Rad51
2-ahela katke
Spo11(meioos), Ho nukleaas
1-ahelalise DNA teke
RecBCD, helikaas/nukleaas
MRX valk (Rad 50, 58, 60
nukleaas)
Ühenduse liikumine
RuvAB
Rad52, Rad53
Ahelavahetuskompleks
RecBCD, RecFOR
Ühenduse lahendamine
RuvC
?Mus81, etc
Koht-spetsiifiline rekombinatsioon ja transpositsioon
:
20
•
Konservatiivne koht-spetsiifiline rekombinatsioon (fosfodiestersidemete
asendamine)
•
Transposoonid e. Hüppavad geenid (DNA vahendatud ja RNA vahendatud)
Et see toimuks on vaja rekombinatsiooni signaaljärjestusi, mida rekombinatsiooni ensüüm ära
tunneks. See järjestus võib ümber pöörduda ehk inversioon või ära kaduda.
Transposooni puhul aga transposeeruv osa liigub ühest molekulist teise.
Remobineeruvat osa saab viia molekuli sisse – insertsioon.
Kui rekombinatsiooni saidid on sama suunalised, siis tekib rekombinatsiooni tulemusena
deletsioon, kui erineva suunalised, siis inversioon (ümber pöördumine).
Rekombinatsiooni saitide vahele jääb rekombineeritav DNA. Türosiin rekombinaarsed ja Seriin
rekombinaarsed. Seriin
rekombinaarsed seostuvad rekombinatsiooni saitidega. Tekitab
mõlemas DNAs kaksikahelalise katke. Iga ahela jaoks rekombinaasi molekul. Tekib kovalente
side, 5’-fosfaat on seotud Seriini hapnikuga.
Rekombinatsioonist võtab osa 4 DNA ahelat. Rekombinatsiooni molekulid seonduvad omavahel.
Pärast rekombinaasi molekul ühendab ahelad omavahel uuesti.
2 tüüpi rekombinaasid: Ser (nt transkriptsioon3 rekombinaas) ja Tyr rekombinaasid (nt faagi P1
Cre rekombinaas).
Kui tekib rekombinatsiooni sünaps – 4 ahelat on kokku saanud, siis tehakse lõige – alles siis
toimub rekombinatsioon. Rekombinatsiooni sünapsis toimub ahelate vahetus.
Ser hapnik atakeerib ühte kindlat fosfaati, tekib fosfoesterside. Niimoodi hoiab rekombinaas
DNAd kinni.
Tyr rekombinaasidel toimub asi üsna sarnaselt. Benseeni tuuma küljes olev
hapnik atakeerib fosfaati, tekib fosfoesterside. Tekib Holliday ühendus.
Kui on kaks erinevat rõngakujulist DNA molekuli, siis tulemuseks on
insertsioon, need
ühendatakse omavahel.
Geeni vahetus homoloogilise rekombinatsiooniga
: üle 50 aluspaari järjestus, mis on
homoloogne algset geeni piiravate järjestustega. Rekombinatsiooniga vahetatakse välja
see osa ning selle uue geeni avaldumise järgi saab selekteerida baktereid, nt
antibiootikumide resistentsus geen.
Homoloogne järjestus peab olema üle 50ne aluspaari (u 200 aluspaari). Kui lisame veel
ühe rekombinaasi molekuli, mis suurendab homoloogilise rekombinatsiooni sagedust.
Geen, mida uurime, on kahe homoloogse järjestuse vahel, mille abil saame selekteerida
baktereid. Nt antibiootikumid (kanamütsiin). Kui antibiootikumi resistentsuse geen
esineb, siis bakterid muutuvad resistentseks selle antibiootikumi suhtes. Kui külvame
plaadile, jäävad resistentsusega bakterid seal elama.
Transgeense hiire valmistamine DNA rekombinatsiooni abil: Tahame uurida mingi kindla geeni
tähtsust, seda vajadusel välja lülitades. Hiire embrüonaalsetesse tüvirakkudesse viime DNA,
mis sisaldab samu järjestusi, sarnane, aga seal on samasuunalised rekombinatsiooni
järjestused, LOX saidid. Meid huvitavad geenid on LOX saitide vahel. Lisaks neil on seal veel
geen, mida kutsutakse neogeeniks, mis annab neomütsiini resistentsuse (nagu kanamütsiin).
Kui selle DNA viime embrüonaalsesse tüvirakkudesse, siis DNAd tahetakse ära parandada,
21
seetõttu toimub rekombinatsioon. Saame rakud, mis on neomütsiini resistentsed. Need
embrüonaalsed tüvirakud süstitakse hiire embrüosse, tekib mosaiikne embrüo. Et seda näeks
paremini kasutatakse erinevat värvi hiireliine. Seega hiirepojad on nii mustad, kirjud kui
valged. Kirjud on kimäänsed, osa tüvirakke on pärit emalt, osad on need muudetud. Kui nüüd
neid omavahel ristata siis osad on mustad, enamus kirjud ja osad valged. Saab tekitada täiesti
valge hiireliini. Lõpuks saame hiire, kus LOX saitide vaheline DNA on deleteeritud.
Cre rekombinaas
: tunneb ära LOX saidi. Koosneb kahest domäänist ja nende vahel olevast
hästi liikuvast elemendist. Võib esineda erinevas konformatsioonis. Aminoterminaalne domään
ja karboksüterminaalne domään. Mõlemad domäänid osalevad DNAga seostumisel.
Karboksüterminaalse domääni juurde moodustub aktiivtsenter, mille küljes on türosiin.
Seostuvad DNAga niimoodi, et ainult üks Cre rekombinaasi dimeeridest on korraga aktiivne.
Mõlemad küll seostuvad korraga DNAga, aga katkemise DNAs põhjustab ainult üks Cre
rekombinaasi molekul, üks reageerib DNAaga teine ei reageeri. Rekombinatsiooni sünaps –
DNA ahelad on risti ja 4 rekombinaasi molekuli on kõik omavahel seotud, millest 2 on aktiivsed.
Rekombinaasi molekulide seostumine DNAga on asümmeetriline. Kui ahelad vahetuvad
muutuvad teised teine rekombinaas aktiivseks.
Lambda faagi integreerumine peremeesraku kromosoomi
: lüsogeenses tsüklis integreerub
peremeesraku molekuli ja ei ilmuta seal ennest mitte kuidagi, integreerumine toimub kindlasse
kohta, seda kutsutakse attB saidiks. Lambda DNA on normaalselt lineaarne kaheahelaline DNA,
integreerides tehakse temast tsirkulaarne vorm.
IHF integration host factor – seondudes
DNAga tekitab DNA struktuuris kõverdumise. Kui ta on seostunud, siis satuvad Lambda
järjetused üksteisele lähemale.
Transposoonid
Mobiilsed geneetilised elemendid, DNA osad, mis on võimelised liikuma ühest kohast teise.
Võivad liikumise käigus paljuneda või lõigatakse ühest kohast välja ja kleebitakse teise kohta.
Inimese puhul 40% mittekodeerivast DNAst on tulnud transposoonidest.
E. coli DNA: IS1 ja IS186 – insertsiooniline järjestus ehk transposooni laadne element.
Pagaripärm: Ty2 – oluliselt suurem, geenide vahelist ala juba rohkem
Puuviljakärbes: rohkem insertsioonilisi elemente, geenide vahelised alad oluliselt
suuremad
Inimene: mobiilsed geneetilised elemendid põhiliselt, geene vähe
Mais: puha transposoonid järjest
Kõige rohkem on DNAd ühe raku kohta salamandritel, 100 korda rohkem DNAd raku kohta kui
inimesel. Liilialistel ka samuti 100 korda rohkem DNAd kui inimesel (0,05% kodeerivat DNAd) –
DNA paradoks – ei tähenda keerukamat isendit, vaid rämpsDNAd on lihtsalt kõvasti rohkem.
DNA transposoonid
:
1.
Bakteri replikatiivsed transposoonid – omased otsmised pöördkordusjärjestused,
sisaldavad transposaasi geene jm geene, ka antibakteriaalsete ainete resistentsuse geene.
Bakteril tekib resistentsus antibiootikumi tõttu just seepärast, et need geenid on
transposoonides. Liikumise mehhanism: kopeeritakse koos replikatsiooniga ja siis nad liiguvad
ühest kohast teise replikatsiooni käigus. Kasutab nt: Faag Mu (bakteriofaag, viirus, kasutab
paljunemiseks sama mehhanismi nagu transposoonid), Tn3
22
2.
Bakterite lõika ja kleebi transposoonid – samad struktuursed iseärasused. Lõigatakse
algsest DNAst välja ja liiguvad uude DNAsse, iseseisvalt neid ei paljundata. Nt Tn5, Tn10,
erinevate bakterite transposoonid.
3.
Eukarüootide transposoonid – pöördkordused raamistavad intronitega geene,
otsmised kordusjärjestused, mis sageli sisaldavad nii otseseid kui pöörduvaid korduvjärjestusi.
Lõika ja kleebi mehhanismiga paljunevad. Esinevad kõigil eukarüootidel. P element, hAT
perekonna elemendid.
RNA vahendatud transposoonid
: DNA pealt sünteesitakse RNA, RNA pealt sünteesitaks uuesti
DNA ja DNA integreerub uuesti kromosoomi, kui ei integreeru läheb kaduma. RNA vahendatud
transposoonid esinevad ainult eukarüootidel. Põhjuseks, miks eukarüootidel on nii palju
rämpsDNAd.
retrospooni nimetus – info liigub RNAlt DNAle, tagurpidi
1.
Viiruse sarnased retrosposoonid – sisaldavad otstes otseseid korduvjärjestusi, pikad
250-600 aluspaari, ka pöördkordusi(lühikesed). Sisaldavad pöördtrankriptaasi sarnaseid geene,
RTase, Int integraas, RnaseH, RNA helikaase, viiruse sarnaseid valke. Otsesed
kordusjärjestused sisaldavad LTR promootorit, transkriptsiooni alustamise piirkondi. Pöörd-
trankriptsiooni teel sünteesitakse esimene DNA ahel, siis teine ka ja siis see DNA integreerub
peremehe genoomi (insertsioon). Levinud laialt, kuid mitte kõige enam. Nt Ty (pärm), Copia
element (Dros).
2.
Poly(A) retrosposoonid – tõelised genoomi parasiidid, kõige rohkem inimesel neid.
Sisaldavad 3’ otsas AT rikast järjestust. PolyA retrospooonid sisaldavad järjestust, mis on
tekkinud PolyA pealt DNA sünteesist. mRNA on ära sünteesitud, siis lisatakse talle PolyA
järjestus, kui see on aga DNAs siis on ta sinna sattunud RNA pealt pöördtrankriptsiooni teel. 5’
otsas mittekodeeriv piirkond ehk UTR, Rna siduv valk ja RTase. Inimese genoomi on mutantsed
tranpsosoonid, nad ei sisalda enam funktsionaalseid RNAd siduvaid valke. Nende pealt ei saa
valku sünteesida, kuid saab määrata nende päritolu. Neil on sisemised promootorid, kus
alguses sünteesitakse üks osa sellest elemendist ja siis sünteesitakse teine osa otsa,
keeratakse ringi. Transkriptsioon sisemiselt promootorilt, märklaud initsieerib pöörd-
transkriptsiooni (endonukleolüütiline). Nt F ja G elem (Dros), LINE ja SINE elemendid, Alu
järjestused.
Transposoon ei eksisteeri iseseisvalt, on peremehe DNAs. Otsmised pööratud
korduvjärjestused. Otsesed kordusjärjestused tekivad transposooni liikumisel ühest kohast teise
(levimise mehhanismi tulemus). Peab sisaldama kas transposaasi või integraasi ja
pöördtranskriptaasi. Pöördtranskriptaas sünteesi RNA alusel DNA ja integraas sisestab DNA
kromosoomi. Sisaldavad ka veel muid geene.
Retroviirused liiguvad ühest organismist teise RNA kujul, nende elutsüklis jooksul toimub RNA
pealt DNA süntees. Vähkkasvajate viirused on ka retroviirused.
Lõika ja kleebi mehhanism
: Otsmised pöördkorduvjärjestused on
transposaasi äratundmise
järjestused, nendega seondub ensüüm transposaas, mis katalüüsib transposooni liikumist.
Märklaud DNA ahelad lõigatakse niimoodi katki, et jääb üleulatuv 3’ ots. Transposaasid
seonduvad oma otsmiste pööratud korduvjärjestusega, nad on vastupidise sümmeetriaga
seondunud, nagu Cre rekombinaasi puhul oli. Selle tulemusena transposaasid interakteeruvad
23
omavahel, transposoos lõigatakse välja, transposaasid lõikavad kumbki oma ahela katki,
transposooni poole jäävad 3’ otsad, kromosoomi poole 5’ otsad. Transposaas ise (enamus
neist) ei ole võimeline seda teist ahelat katki lõikama. Transposaas ei lõika enne ahelat katki,
kui ei ole olemas kohta teises DNAs kuhu teda panna. Transposooni otsad võidakse tsükliliseks
muuta. Kaheahelaline ots tuleb ära peita, et ei hakataks parandama DNAd. Transposooni otsad
ühendatakse märklaud DNAga nii, et kahe katkemise vahele jääb 2+9 nukleotiidi unikaalset
DNAd. Ühendatakse vastavad ostad.
Replikatiivsed transposoonid
: algab samuti transposooni tekkimisega, transposaasid omavahel
interakteeruvad, DNA ahelad tuuakse omavahel kokku. Tekivad ainult vabad 3’ otsad
transposoonil, ei lõigata teda välja DNA ahelast. Toimub replikatsioon, transposooni rakuline
polümeraas paljundab transposooni mõlemad ahelad, tekib uus koopia tranposooni.
Kui transposoon oma koha pealt kunagi lahkub, jääb maha transposooni jälg. Kui transposoon
läheb valgu geeni sisse, siis see valk enam ei tööta. Kui transposoon välja lõigata, siis ikka valk
ei hakka uuesti tööle, sest sinna jääb transposooni jälg, rikub ära lugemisraami, kui ta just
mingi ülipisike ei ole, nt mingi 9 aluspaari.
Bakterites ei paljundata transposoone nii mõttetult palju kui ekarüootides. See on sageli
reguleeritud RNA kaudu. Transposaasi geenil on kaks promootorit. Sisse poole ja välja poole
suunatud promootorid. Kui tekib kaheahelaline mRNA, siis see alati lagundatakse, RNA viiruste
vastane kaitsemehhanism. Tn4 perekonna transposaasi aktiivusus surutakse alla tänu sellel, et
tekib kaheahelaline RNA, mis kohe lagundatakse.
Immuunoglobuliinide geenid ja nende rekombinatsioon
:
•
Imetaja (ka inimene) on võimeline tootma 1 000 000 erinevat antikeha, millest
igal ühel peab olema oma geen
•
Immuunoglobuliinide geene on aga vaid mõni tuhat
•
Valgud sünteesitakse alati N otsast C otsa suunas, N-ots on algus
Immuunvastus on omane kõigile elusorganismidele, võib olla erinev.
Rakuline- ja
humoraalne immuunvastus. Humoraalne põhineb
antikehadel. Imetaja on
võimeline tootma miljon erinevat antikeha, millest igal ühel peab olema oma geen. Antikehade
geenid muutuvad elu jooksul. 21 888 valgu geeni inimesel, miljonit ei ole, neid tuleb tekitada.
Immuunoglobuliinide geene on inimesel kõige rohkem, neid on siiski ainult mõni tuhat. Elu
jooksul tekib neid juurde ja võib olla kuni miljon, aga mitte kõigis rakkudes. Immuunoglobuliini
geenide arv muutub erinevates rakkudes erinevalt elu jooksul.
Üks geen kodeerib ühte produkti, immuunoglobuliini puhul aga üks geen kodeerib osa ühest
produktist.
Immuunoglobuliin koosneb
kahest raskest ja kahest kergest ahelast, identsed omavahel. Antigeeni ära tundvad otsad moodustuvad raske ja kerge ahela alguses, N-
ots on algus.
Kõik immuunoglobuliinid on erinevalt ülesehitatud, kuid kõik koosnevad rasketest ja kergetest
ahelatest ja antigeeni ära tundmise piirkond on N-terminaalses otsas raskel ja kergel ahelal.
Raske ja kerge ahel on omavahel kokku liidetud
S-S sildadega, aminohape Cys vahel
kovalentne side, mida on võimalik kergesti tekitada ja kergesti lahti harutada, natuke H-sideme
moodi, aga stabiilsem. S-S sidemed määravad immuunoglobuliini struktuuri. IgA on
24
sekreteeritav valk, moodustub niimoodi, et lisaks
tavalistele konstantsetele ahelatele veel J-ahel ja
sekreteeritav komponent.
Inimese antikehad: IgM, IgD, IdG, IgA, IgE
(allergiaga seotud)
Rasked ahelad on kõigil erinevad (μ,δ,γ,α,ε).
Kerged ahelad on kõikidel täpselt sama tüüpi, neid
on
kahte klassi.
IgG koosneb korduvatest struktuursetest üksustest. Kergel ahelal on üks domään N-
terminaalses otsas. Variaabel piirkonnas moodustub antikeha ära tundev osa. Kerge ahel
koosneb kahest struktuursest üksusest – immuunoglobuliini domään. Raske ahel neljast
domäänist. Mõlemal ahelal kõige N-terminaalsem osa moodustab antikeha ära tundva
piirkonna. Kerged ahelad on rasketega ja rasked omavahel seotud S-S sildadega. N-
terminaalsed domäänid on variaabel piirkonnas, ülejäänud on konstantsed osad.
Iga immuunoglobuliini domään koosneb beeta-tünnikesest, mis koosneb antiparaleelsetest
beeta-lehtedest. Keskel on tühi koht. Beeta-lehtede otstes olevad järjestused moodustavad
selle osa, mis antikeha ära tunnevad.
Lambda, kappa, raske ahela geeni osi inimesel kõiki 300. Hiirel aga raske ahela geeni osi üle
tuhande, kappa omasid 1000 kanti, lambda omasid 2 ainult. LAmbda ja kappa kerge ahela
geeni segemendid. Need on variaabli omad. Konstantseid segmente on kõvasti vähem.
25
Lambda perekonna kergeahela variaabli ja konstantse osa kokkupanek: tulemus-
immuunoglobuliini kerge ahel koosneb variaabel osast ja konstantsest osast. Selline valk
tranlatseeritakse kindla mRNA pealt. Kuidas see mRNA saadakse? Võtame tavalises keharakus
olevad immuunoglobuliini geenid, variaabel ja konstantse geeni segmendid paiknevad erinevas
kohas. Variaabel segment ühendatakse konstantse osaga somaatilise rekombinatsiooni käigus.
(Rekominatsioon, mis toimub somaatilistes rakkudes väga kindlate reeglite järgi). Üks variaabel
valitakse välja ja tuuakse konstantse osa lähedale. Variaabel geen koosneb liiderjärjestuse
osast(ei sünteesita sellelt valku), intronist(lõigatakse välja täpselt nukleotiidi pealt mRNA
tasemel splaissinguga) ja variaabli domääni kodeeriv osa. Konstantne geen koosneb J-
segmendist ja konstanset domääni kodeerivast geenist, nende vahel ka intron.
Kappa perekonna… : Variaabel osa sama nagu lambda perekonna omal, koosneb
liiderjärjestus, intron ja variaabli domääni kodeeriv osa. Konstantne domään sisaldab palju
erinevaid J-segmente. Kahte tüüpi rekombinatsioon. Esimene siis kui variaabel osa ühendatakse
V-J segmendiga, ühendatakse väga täpselt Olenevalt, millise J-segemendi ette jäi variaabel osa,
see jääb alles, kõik teised J-segemendid eemaldatakse splaissimise teel. Teine toimub
splaissimise tasemel.
S.Tonegawa avastas V-J ühenduse.
W.Gilbert nägi, et geeni segmendid vastavad valgu struktuursetele üksustele. Tegelt on see
nii ainult immuunoglobuliinidel.
Raske ahel esimese somaatilise rekombinatsiooni käigus tekib D-J ühendus. Variaabli osa
tuleb tuua D osa lähedale ja siis V-D ühendus. Intronid eemaldatakse splaissimisega.
RAG1 ja RAG2 vastutavad selles eest, et somaatiline rekombinatsioon toimub juhuslikult, kuid
ühendamine toimuks nukleotiidilise täpsusega. Variaabel segemendi lõpus ja J-segmendi
alguses on kindel ära tundmise järjestus.
Somaatilise rekombinatsiooni mehhanism
: ära tundmise signaali juurde tehakse mõlemasse
ahelasse katke, jääb 3’ vaba ots, tekib üleulatuv 5’ ots, need mõlemad otsad peidetakse ära,
tekitatakse juuksenõela struktuur, 5’ ja 3’ ots ühendatakse. Vahepealsel osal on kaks
kaheahelalist katkestust, see tsükliseeritakse, lõpuks see osa lagundatakse. Jääb nüüd alles
osa, kus variaabel osa on ühendatud J-osaga. Funktsionaalne geen tekib.
Somaatilise rekombinatsiooni teel saadakse primaarne immuunvastus, seda tugevdatakse
somaatiliste mutatsioonide teel. Neid tekitab DNA pol∏. Ta replitseerib DNAst ainult
liiderjärjestus, variaabel ja J-osa. Tekitab lisa mutatsioone, mille hulgast on võimalik välja valida
klonaalse rekombinatsiooni käigus vajalik.
T-DNA: bakterist üle kantav DNA
Kaheidulised kõik taimed, mis ei ole teraviljad.
Bakteri DNA ja taime DNA võtavad osa rekombinatsioonist. Bakter geenid integreeritakse taime
genoomi, see mõjutab taime kasvu. Transgeensete taimede tegemisel kasutatakse seda.
Põhineb samuti somaatilisel rekombinatsioonil, aga täiesti erinevat päritolu rakud.
Agrobakteeriumil võime süstida oma DNAd taime rakkudesse. Tal on peale kromosoomi ka
diplasmiidid, selles üks kindel osa kantakse üle taimerakku, kus ta läheb taime kromosoomi.
Selle peale T-DNAst areneb välja kasvaja, mini taim, millel on oma võrsed, lehe, juur.
26
T-DNA koosneb kindlatest geenidest: virulentsuse geenid, võrse kasvu ja juure kasvu
indutseerivad geenid. Nopaliini või oktopiinid kutsuvad esile kasvaja teket. Ja ka veel need
geenid, mis on vajalikud bakteris kasvamiseks, paljunemiseks.
Bakteri poolne DNA on seotud korduvjärjestustega, mis raamistavad üle kantavat DNAd.
Taimerakus ta integreerub mitte spetsiifilise otste ühendamise teed pidi, pole kindlat kohta,
kuhu integreerub. Vasak ots pole väga täpselt määratud, parem aga on.
Transkriptsioon
RNA koostises riboos. RNA stabiilsus ja ruumiline struktuur on oluliselt erinev võrreldes DNAga.
RNA sünteesi viib läbi DNA sõltuv RNA polümeraas. Protsessi nimi transkriptsioon. RNA
sünteesitakse ainult ühe DNA ahela pealt. Üks on
matriits ahel, teine
kodeeriv ahel.
Kodeeriv ahel on täpselt sama järjestusega nagu sünteesitav RNA.
RNA polümeraas seondub DNAl mingi kindla piirkonnaga. Alustab sünteesi, sünteesib kuni
terminatsiooni signaalini, RNA vabaneb. RNA polümeraas viib läbi RNA sünteesi. Neid on kahte
tüüpi. Me vaatame neid, mis koosnevad paljudest polüpeptiididest. Struktuurilt DNA
polümeraasi sarnased, aga koosnevad paljudest polüpetiididest erinevalt DNA polümeraasist.
Konserveerunud. RNA polümeraasi võrreldakse
vähi sõraga. Vao põhjas asub aktiivtsenter.
Nagu DNA puhul matriitsahel, mille järgi sünteesitakse, aktiivtsentris toimub substraadi välja
valimine. RNA polümeraas tunneb spetsiifiliselt ära 2’ hapnikke, mida desokünukleotiididel
pole. RNAsse lülitatav nukleotiid valitakse välja komplementaarsuse alusel, geomeetrilise
sobivuse alusel, peab täpselt ära mahtuma aktiivtsentrisse, H-sidemetega paardumine peab
olema. Kaks kahevalentset metalliiooni, mis üks osaleb desoküaniooni moodustamisel, prooton
eemaldatakse ja see reageerib järgmise ribonukleotiidi alfa fosfaadiga.
Kolm etappi:
1.
RNA polümeraas seondub kindla
piirkonnaga DNAl – geeni promootoriga. RNA
polümeeride seondumine DNAga ja 10-12 esimeste
nukleotiidide süntees on transkriptsiooni
initsiatsioon (alustamine). Selle käigus RNA
polümeraasi konfirmatsioon muutub, läheb üle
protsessiivsesse konformatsiooni ehk ei lase RNA
polümeraasil DNA ahela küljest lahti tulla.
2.
Ahela pikendamine –
elongatsioon
3.
Terminatsioon – RNA vabaneb
Prokarüootide puhul on kõige tähtsam promootori
ära tundmisel sigma faktor, määrab ära millist
ahelat hakatakse transkribeerima ja kust kohast
hakatakse sünteesima.
1.
RNA polümeraas koos sigma faktoriga seondub DNA ahelaga, tekib
suletud kompleks
(DNA on kaheahelaline).
27
2.
Avatud kompleksi teke, kõikide komponentide ruumiline struktuur muutub. DNA ahel
harutatakse lahti. Sigma faktor muudab ka konfirmatsiooni, RNA polümeraas ka. Tekib
transkriptsiooni mull – DNA ahelad on omavahel lahutatud seal.
3.
Terminatsiooni signaalid on küll kirjas DNA peal, kuid neid seal ära ei tunta, alles RNA
tasemel tuntakse.
Promootori puhastamine – initsiatsioonilt üleminek elongatsioonile.
Initiatsioon eeltuumsetel
:
Sigma-faktor tunneb ära promootori(-10, -35 boxid). Siis korraldab RNA
polümeraasi nii, et süntees algab õigelt ahelalt(matriitsahelalt). Seda teist
ahelat, mida ei kasutata kutsutakse kodeerivaks ahelaks, aga ta on ju
identne sünteesitava RNA-ga.
Katalüüsib ka RNA polümeraasi ja DNA vahelise dünaamika kujunemist.
Spetsiifiline seondumine on tunduvalt tugevam mittespetsiifilisest.
Esialgne etapp, seondumine on väga kiire.
Mulli tekkimine ka kiire, kuid veidi aeglasem. Kinnine kompleks avatud
kompleks.
Sünteesi alustamine ise toimub kiiresti.
Limiteeriv etapp on promootori puhastamine st initsiatsiooni üleminek
elongatsiooniks.
Elongatsiooni kiirus – 40-80 nukl/sek. DNA süntees umbes 1000/sek. Take
that. Põhjuseks see, et RNA ja DNA heterodupleks nõuab lahutamist, mis
nõuab ise energiat.
Abortiivne initsiatsioon – promootorilt toimub pisikeste RNA juppide süntees.
DNA sünteesi puhul lahutavad ahelaid helikaasid, RNA polümeraas lahutab ahelaid aga ise. RNA
sünteesi käigus produkt vabaneb ja RNA dissotseerub RNA kompleksist. RNA polümeraas
lahutab DNA ja RNA ahelad ning aitab DNA ahelatel taas kokku sulanduda, lükkab eemale RNA.
Kui lülitub ahelasse vale nukleotiid, siis RNA polümeraas käigu pealt parandab vea ära. Jääb
korraks seisma, lõikab vale nukleotiidi välja ja läheb sünteesiga edasi. RNA polümeraas
sünteesib ja lagundab.
Pürofosforolüütiline parandamine – kui viga on viimases nukleotiidis. Kui aga viga ei ole
viimases nukleotiidis vaid eespool, siis RNA polümeraas jääb seisma ja lagundab ära kuni
vigase piirkonnani.
Elongiinid – aitavad tõsta RNA polümeraasi protsessiivsust. Vigade parandamise käigus võib
RNA dissotseeruda RNA polümeraasi küljest, et see ei juhtuks, ongi vaja elongatsiooni faktoreid,
nt Gre faktor.
Transkriptsiooni terminatsiooni on kahte tüüpi: ühed ei vaja lisafaktoreid – iseeneslik, teine on
rho-sõltuv terminatsioon, vajab rho-faktorit.
Rho-sõltumatu
tel järjestustel on pööratud kordusjärjestused. Sünteesitav RNA on
komplementaarne iseendaga, paardub iseendaga. Kaheahelalise RNA piirkonnale järgneb A-T
rikas ala. Kaheahelaline RNA lükkab enda DNA küljest lahti, sest RNA-RNA kompleks on
28
stabiilsem kui RNA-DNA kompleks. AT paarid, mis järgnevad on vähem stabiilsed ja selle koha
pealt tuleb RNA DNA pealt lahti ja RNA polümeraas ei saa enam sünteesi jätkata. Niimoodi
toimibki rho-sõltumatu terminaator.
Rho-sõltuv
al osaleb helikaas (rõngas ümber RNA), mis seondub eelistatult RNA 5’ otsaga ja
liigub nagu RNA polümeraasiga kaasa. Kui RNA polümeraas kohtab oma teel rho-sõltuvat
faktorit, jääb RNA polümeraas seisma, seondub profaktoriga. RNA ahel lõigatakse katki RNA
polümeraasi küljest. Ongi terminatsioon.
Sama aegselt sama geeni saavad transkribeerida mitmed erinevad RNA polümeraasid. DNA
replikatsioonil ei saa nii. RNA polümeraas ei vaja sünteesi alustamiseks praimerit. Bakterites
see sama RNA polümeraas, mis sünteesib RNAd sünteesib ka replikatsiooni praimereid.
Eukarüootidel on kolm erinevat RNA polümeraasi, need erinevad nii subühikute poolest, milles
koosnevad, kui ka selle koha pealt, milliseid geene nad transkribeerivad.
On ka ühe subühikulisi RNA polümeraase, mis on struktuurselt sarnased DNA polümeraasidel,
esinevad enamasti bakteriofaagidel. Suudavad siiski alustada RNA sünteesi ilma praimerita,
kuid on DNA polümeraasi sarnane.
RNA polümeraas
Beeta’ subühik sisaldab katalüütilist tsentrit. Beeta ja beeta’ vahel auk, kust läheb läbi/tuleb
välja RNA. 2 alfa subühikut, mis moodustavad dimeeri. Oomega subühik. Sigma subühik on
haraline moodustis, mis seondub beeta ja beeta’ subühikute vahele, põhiliselt beeta’
subühikuga. Kompleksis RNA polümeraasiga on sigma subühik selline, mis koosneb 2st
domäänist (teineteisest võrdlemisi kaugel), koosneb ainult alfa heeliksitest. Kui sigma faktor ei
ole kompleksis RNA polümeraasiga, siis tema struktuur ei ole defineeritav. Kahevalentsed
metalliioonid, eriti Mg ioonid on RNA struktuuri moodustumisel olulised. RNA polümeraasil on
mitmeid Mg ioonide sidumise kohti.
Promootor
Transkriptsiooni alguspunkti tähistatakse +1ga. RNA ahelad algavad reeglina puriiniga A või G.
Esimesest sünteesitavast nukleotiidist ette poole jäävat osa nimetatakse ülevoolupiirkonnaks.
Taha poole jäävat osa allavoolu piirkonnaks. 10 ja 35 aluspaari eespool on sigma faktori
äratundmise piirkonnad(-10, -35 box), millega seonduvad sigma faktori kaks domääni. UP-
elemendi nimelise järjestusega on tugevamad promootorid, sellega seondub RNA polümeraasi
alfasubühik. On ka sellised, millal on ainult 10 juures lisa järjestus, 35 ja UP pole. 10 koha peal
toimub ahelate lahutamine, trankriptsiooni mull moodustub sinna, AT rikkas koht.
+ ahel ehk kodeeriv ahel on see, mis on RNA ahelaga identse järjestusega, selle pealt ei
sünteesita RNAd. Komplementaarne ahel on see mille pealt RNA süntees toimub.
Transkriptsiooni terminatsioon
:
Rho sõltuv terminatsioon vajab RNA järjestuse signaali ja rho faktorit. Rho faktor liigub 5’-3’
suunas. Kui saab RNA polümeraasiga kokku (liigub talle järgi, RNA polümeraas jääb seisma),
siis transkriptsioon lõppeb.
Rho sõltumatu terminatsioon ei vaja lisafakroteid, RNA signaal koosneb pööratud kordustest,
mis moodustavad kaksikahela (juuksenõela) ja selle järgneb U-rikas järjestus.
29
Antiterminatsioon
:
Trankriptsiooni antiterminatsiooni faktorid seonduvad sünteesitava RNA’ga ja RNAP’ga ning ei
lase rho faktoril toimida, ei lõppe transkriptsioon.
Bakteriofaagidel on eriti head antiterminatsioonifaktorid. Bakterites ka olemas ja eukarüootides
ka.
Antiterminatsiooni faktorid on valgud, mis seonduvad nii RNA kui RNA polümeraasiga.
Seostuvad RNA 5’ otsaga ja seondavad RNA ja RNA polümeraasi. Normaalne rho faktor seostub
RNA 5’ otsaga, kui see ots on aga valkudega kompleksis, ei saa ta ühineda RNAga, ei jõua
järele RNA polümeraasile. Nus A, Nus B (C,D,E) jne faktorid tõstavad transkriptsiooni
protsessiivsust, ei lase rho sõltuval terminatsioonil toimida. Mõjutavad ka tekkiva RNA
sekundaarstruktuuri tekkimise kiirust. +
On ärahoidmaks juhusliku terminatsiooni juhtumisel, eriti rRNA sünteesidel
esinev.
Et suurendada RNA sünteesi protsessiivsust on olemas antiterminatsiooni
faktorid.
Need faktorid on valgud, mis seonduvad RNA polümeraasiga ning
muudavad ta rokem protsessiivseks, aga nad paljudel juhtudel lisaks ka
seonduvad RNA 5’ otsaga, kui see tuleb tunnelist läbi ning hoiavad seda 5’
otsa kogu aeg polümeraasi küljes. Nüüd see kasvav RNA läheb aina
pikeneva linguna. Selleks vajalik, et heksameerne Rho faktor seondub
kergesti RNA 5’ otsaga. Aga antud juhul ei saa ta sinna seonduda ning
sellega ei suuna ka trankriptsiooni terminatsiooni.
See interaktsioon RNA 5’ otsa ja sünteesitava ülejäänud ahelaga paneb
otsad lähestikku.
Mis on geen
:
Pärilik teave säilitatakse nukleiinhapetes (DNA, RNA) nukleotiidse järjestusena. Geen on DNA
(või RNA) lõik, mis kodeerib ühte eralduvat produkti. Geenid võivad kattuda nii ühel ahelal kui
eri ahelail. Ühe geeni sees võib asuda ka teine geen. Geen on pigem informatsioonikandja,
informatsiooniline element.
Geenide kattuvus on omane just viirustele, bakteriofaagidele, sest neile on tähtis, et genoom
oleks väga kompaktne ja kiiresti paljundatav.
Eukarüootide geenidele on omased intronid ja eksonid. Intron kodeeritud RNAd on
snoRNAd,
nad on vajalikud teiste RNA molekulide protsessimiseks ja modifitseerimiseks.
Erinevad geenid
„Koduhoidjad“ geenid – vajalikud kõikides rakkudes, alati ühte moodi, struktuursete
komponentide ja põhiainevahetuse läbiviimiseks vajalikud, nt DNA polümeraas, RNA
polümeraas, ribosoomi komponendid, histoonid, helikaasid, topoisomeraasid jms. | Kodeerivad
valgu või RNA molekule, mis on vajalikud põhiliste eluprotsesside läbiviimiseks(nukleiinhapte ja
valkude metabolism, põhiainevahetus, raku struktuurse komponendid). Nad avalduvad kõigis
kasvates rakkudes.
30
Indutseeritavad geenid – neid pole koguaeg vaja. Mõnes koes ekspresseeruvad ühed geenid,
teistes teised. Või mingites kindlates elutingimustest ekspresseeruvad mingid kindlad geenid. |
Indutseeritavad geenid avalduvad vaid kindlates tingimustes kui nende produktid on vajalikud.
Nende avaldumiseks vaja induktor ehk tingimused
RNA geenid – geeni produkt on RNA, nt rRNA, tRNA, väiksesed RNAd
Valgu geenid – geeni produkt on valk, nt mRNA geenid
Regulaatorid, nt promootor ja pseudogeenid ei ole üldse geenid (produkti mõttes)
Pseudogeenid
– need ei kodeeri funktsionaalset produkti. Neil kas terminatsioon toimub liiga vara,
ei sünteesi tervet produkti. Või sünteesib ta valgu geenina ka mitte terve valgu ahela või
puuduvad tal RNA protsessimiseks vajalikud signaalid.
Geeni aktiivsuse regulatsioon
: On reguleeritud mitmel erineval tasemel.
1.
Trankriptsioon – RNA sünteesi reguleerivad valgud ja neid mõjutavad faktorid. Kus ja
millal toimub, kui palju, jne.
2.
RNA protsessimine ja transport – bakteritel suhteliselt vähe oluline, eukarüootidel
aga väga oluline
3.
mRNA lagundamine (eluiga) – võib olla väga lühike kui ka väga pikk
4.
Translatsioon – kui palju RNA pealt sünteesitakse valku. mRNA struktuuri, ribolülitite,
regulaator valkude ja RNAde kaudu toimiv regulatsioon.
5.
Valgu modifitseerimine ja lokalisatsioon – bakterite puhul vähe tähtis
6.
Valgu eluiga – bakterite puhul olulisem, nt DnaA valk saa osaleda ainult üks kord
replikatsiooni initsiatsioonil.
Prokarüootidel transkriptsiooni regulatsioon
Transkriptsioon ja translatsioon on omavahel seotud, toimub samal ajal. Väga oluline geeni
aktiivsuse regulatsiooni tase on ribosoomide süntees, eukarüootides see ei ole, sest ribsoomide
arv ei muutu nii palju.
Kõige olulisem geeni regulatsiooni alus on valgu ja nukleiinhappe vaheline spetsiifiline ära
tundmine. Kui valk seob trüptofaani, siis see indutseerib valgus konformatsiooni muutuse nii, et
mõlemad alfa-heeliksid saavad samaaegselt ühineda DNA kindla järjestusega, oma ära
tundmise piirkonnaga.
Geeni regulaatorid on enamasti kahe domäänsed valgud, N-terminaalne domään seostud
DNAga. Need DNAga seonduvad piirkonnad tunnevad ära oma kindlad DNA järjestused ja
seonduvad sellega, C-terminaalsed järjestused tüüpiliselt interakteeruvad omavahel. Üks faktor
tavaliselt ei saa ise seonduda DNAga tugevalt, kui neid on aga kaks, siis nad abistavad üksteist
ja saavad seonduda tugevalt, moodustavad koos mingi kindla struktuuri. Valgu alfa-heeliksis
olevad aminohapped tunnevad ära kindlad järjestused DNAl.
Transkriptsiooni initsiatsioon, regulatsioon
Algab
DNA kindlas piirkonnas – promootoris. Sellega seondub RNA
polümeraas. Sigma-faktor on abiline siin, mis tunneb promootori ära
31
tunneb. Promootorid on alati ebasümmeetrilised järjestused, mitte
pööratud kordused.
tekib suletud kompleks, siis avatud kompleks. Siis hakkab transkriptsiooni
alustamine. Ajaliselt limiteeriv etapp siin promootori puhastamine, see on
kui RNA pol läheb initsiatsiooni faasist elongatsiooni faasi.
Reaalselt on harva leida ideaalseid promootoreid. Promootorid on
enamjaolt võrdlemisi nõrgad. Selleks et transkriptsioon saaks alata on vaja
faktoreid tegemaks promootoreid tugevamaks. See on üks
geeniregulatsiooni aluseid.
Aktivaator seondub aktivaatorjärjestusele, activator binding site, mis on
promooteri ees. See seostumine tõstab transkriptsiooni initsiatsioonitaset.
Aktivaator seondub ühelt poolt DNA-ga teiselt poolt polümeraasiga.
Seondumine on selle abil stabiilsem ja sagedasem. Iga seondumine on
produktiivne ja vib sünteesini, ilma see nõnda alati pole
Effekt on allosteeriline. Ilma aktivaatorita on avatud kompleksi teke
oluliselt aeglasem. Aktivaator muudab polümeraasi struktuurset
üleminekut, mis peab toimuma avatud kompleksi tekkimise juures. Peab
muutuma nii polümeraasi kui ka DNA struktuur. Aktivaator hõlbustabki
seda.
Repressoril on vastupidine efekt. Ta seostub operaatorile, see on
promooteris. Operaator on -10 ja -35 bokside vahel.
Ta takistab RNA polümeraasi ja sigma-faktori kompleksi moodustumist. Ei
lase lihtsalt DNA-le ligi.
Samamoodi võib repressor takistada ka avatud kompleksi teket, antud
juhul on repressori seondumispiirkond promooteri ees.
Lambda repressori seondumine: Tema DNAga seondumise struktuur on erinev alfa-heeliksist,
koosneb heeliksitest ja pööretest. Tal on küll heeliksid, aga need ei seondu DNAga otseselt.
DNAd ära tundvad alad peavad olema kindlal kaugusel üksteisest ja kindla orientatsiooniga,
sest muidu ta ei saa DNA järjestusega seonduda.
Transkriptsioonifaktoreid jaotatakse selle järgi, millise struktuuriga nad ühinevad DNAga.
Transkriptsiooni negatiivne kontroll
: põhiline bakterite maailmas, kui regulaatorit ei ole on
geen aktiivne. Kui reguleeritakse, siis surutakse maha geeni aktiivsust. DNA ei ole nii tihedalt
seotud, RNA polümeraas saab DNAga seonduda ja transkriptsiooni alustada.
Bakterite geenid on organiseeritud
operonidesse, st ühe promootori järel on mitu erinevat
geeni. Kui DNA on vaba, valke taga seotud pole, saab RNA polümeraas promootoriga vabalt
seonduda, toimub transkriptsioon. Kui aga promootori lähedusse - operaatori piirkonda –
seondub regulaator valk, siis RNA polümeraas ei saa promootorile seonduda, sest regulaator on
seal juba ees. Need geenid on välja lülitatud. Regulaatorvalke kutsutakse
repressoriteks.
Positiivne kontroll
: omane eelkõige eukarüootidele, kui geeni ei reguleerita, siis ta on
inaktiivne. DNA on väga tihedalt kokku pakitud kromosoomidesse ja see ei ole kätte saadav
RNA polümeraasile. Eukarüootide geenid on pakitud nukeleosoomidesse, regulaatorvalgud neid
üle ei leia. Et transkriptsioon saaks toimida, tuleb kõigepealt kromosoom lahti harutada ja siis
toimub geeni aktivatsioon kui regulaatorvalgud on DNAga seostunud. Regulaatorvalke
kutsutakse
aktivaatoriteks.
32
Lac operon – Avastasid F. Jacob ja J. Moned. lacZ, lacY ja lacA geenidest koosneb.
lacZ on
ensüüm beeta-galaktosidaas, kasutatakse eukarüootide uurimisel selle geeni avaldumise
kaudu, rakud lähevad tumesiniseks kui ensüüm toimib.
lacY geeni produkt on permeaase –
valk, mis võimaldab galaktoosi, ka laktoosi sisenemist rakku, permeabiliseerib raku.
lacA produkt on transatsetülaas, ka suhkruga seotud.
lacI produkt on regulaatorvalk, repressor, kui
ta on DNAga seotud, siis ta lülitab transkriptsiooni välja.
Kui organismis laktoos, siis selle bakteri lacI repressoriga seonduvad
laktoosi molekulid, mis selle desaktiveerivad ning saab toimuda vastaval
bakteril operoni aktivatsioon, nüüd saab toimuda valgu süntees mRNA-lt.
Kogu keskkonnas olev laktoos tarbitakse ära, induktorit pole, siis lacI
repressor taaskord seondub DNA-ga. Ja ei toimu vastavate mRNA-de
süntees ja seega ka vastavate valkude süntees. mRNA eluiga on lühike,
see lagundatakse, valkude oma seevastu on pikk.
Kui on kas lacI geen mutantne või on lacI geeni seondumispiirkond
mutante, siis muutub lacI operon konstitutiivselt aktiivseks.
Operaatorpiirkond geenis on repressori sidumiskoht.
mRNA on suhteliselt lühikese elueaga molekul. Kui süntees indutseeritakse, siis sünteesitakse
mRNA, kui induktor ära kaob, mRNAd juurde ei sünteesita ja lagundatakse ära mRNA. Eluiga on
minutites, natuke rohkem kui 10 minutit on ta juba kadunud. Valk aga jääb alles, kui
sünteesitakse, ei kao kuhugi.
Lisaks repressoritele on olemas ka aktivaator valgud, mis võimendavad transkriptsiooni.
Bakterid ei kasuta teisi suhkruid nii kaua kuni on ümbritsevas keskkonnas glükoosi.
CAP valgu (aktivaator) seondumine promootor piirkonna juurde soodustab transkriptsiooni initsiatsiooni. |
CAP valk – kataboliitne aktivaatorvalk. See võib ka seonduda promootorpiirkonda. Võib
takistada repressori seondumist DNA-ga või võib ka stabiliseerida RNA polümeraasi seondumist
DNA-le.
33
Guanosiintetrafosfaat või guanosiinpentafosfaat on regulatoorne molekul, mis mõjutab RNA
polümeraasi aktiivusust. Ta sünteesitakse GTPst ja ATPst, seda sünteesib
gunaosiintertafosfaadi süntetaas ehk ppGpp („võlutäpikese“ süntetaas).
Võlutäpikese süntetaas seostub ribosoomidega, mRNAd aga pole, sellega seonduda ei saa,
hakkab sünteesima guanosiindeltafosfaati, mis seondub RNA polümeraasiga täpselt samasse
kohta, kuhu seondub esimene sünteesitav nukleotiid. RNA polümeraas ei saa aru, et tal juba on
fosfaat 5’ otsas ja 3’ otsas (fosfodiester side oleks nagu juba olemas). Ja blokeerib RNA
polümeraasi, siis kui tal pole vaja RNAd sünteesida. rRNA promootorid on kõige tundlikumad,
need lülitatakse kõige enne välja.
Võlutäpike kuulub alarmoonide hulka. Võlutäpikese süntetaas
ehk
RelA valk sünteesib ainult sel juhul, kui ribosoomide A
sait on tühi, ei jätku aminohappeid või energiat. Nälja tõttu
sünteesitud
ppGpp seondub RNA polümeraasiga ja takistab
RNA sünteesi. ppGpp peatab rRNA(ribosoomide) sünteesi.
Neid sünteesivad ka patogeensed bakterin nt Bacillus subtilis
Alarmoonid on stressi vastus:
ppGpp tase tõuseb mitmesuguste sttressi tingimuste krral
patogeenidel indutseeritakse ppGpp taseme tõus peremehe
immuunvastuse puhul
stressi puhul sünteesitakse ka 6S RNA, mis on RNA polümeraasi
inhibiitor Võlutäpikest sünteesitaks siis, kui rakk jääb nälga.
Blokeerib ära ka RNA polümeraasi.
cAMP muudab CAP valgu struktuuri ja on allosteeriline regulaator. cAMP tase rakus sõltub
glükoosist, cAMPi sünteesitakse siis kui glükoosi tase langeb. Kui cAMP seostub CAP valguga,
siis monomeeride omavaheline orientatsioon muutub ja selle tulemusena heeliksid, mis on
võimelised seonduma DNAga keeravad ennast sellisesse orientatsiooni, et saavad mõlemad
DNAga seostuda.
Attenuatsioon
i klassikaline näide on AH sünteesi operonid, mis lülitatakse välja, kui vastavat
AHd on piisavalt. Bakteritel toimub see, sest transkriptsioon ja translatsioon on omavahel
soetud. Translatsioon mõjutab ka transkriptsiooni. Trp operoni kontrollitakse attenuaatori abil
tänu transkriptsiooni ja translatsiooni seotusele
o Attenuaator – RNA tasemel. On tähtis mehhanism AH-de sünteesi
juures, on tähtis ka mujal. Nt trüptofaani hulk on liiga suur rakus, siis
selle biosüntees lõpetatakse.
o Sünteesitav RNA sisaldab liiderjärjestust, millelt sünteesitakse
liiderpeptiid(mõnikümmend ah-d, tal puudud eriline fn, kas ta sünt
või mitte). Tema süntees määrab ära struktuurgeenide(Trp sünteesi
geenide) ekspressiooni. Effektor on aga see, kas translatsioon
toimub või mitte. Tekib terminatsiooni juuksenõel(rho sõltumatu
term.).
Kui trüptofaani pole, siis ei saa sünteesida liiderpeptiidi, sest
liiderpeptiid sisaldab mitut trüptofaani koodonit. Siis jääb ribosoom
mingi koha peal(Trp koodoni kohas A saidis) lihstalt seisma ning
terminatsiooni juuksenõela ei teki.
34
o Ehk kui rakus kõrge trüptofaani tase, siis lülitatakse välja
transkriptsioon ja see toimub RNA sekundaarstruktuuri kaudu. Sest
siis ei teki hairpin struktuuri, vaid üheahelaline, mis aga pole
terminaatorstruktuur mRNA-l.
o Kui toimub transkriptsioon ja RNA polümeraasi küljest läbi kanali
vabaneb RNA, siis seda mRNA-d hakatakse kohe transleerima.
Valkude süntees stimuleerib omakorda RNA polümeraasi. Kui
translatsiooni ei toimu ei toimu ka stimulatsiooni sellist.
o Liiderjärjestuse mittetranskriptsiooniga avaneb järgmise geeni
startkoodon.
uORF - enne struktuurseid geene on üks pisikene ülesvoolu avatud lugemisraam, mille pealt
saab sünteesida pisikese peptiidi. Nii prokarüootidel kui ka eukarüootidel esineb neid.
Kui trüptofaani pole, ei saa ribosoom translatseerida, ribosoom jääb seisma ja terminatsiooni
nõela ei teki. Kui aga trüptofaani on piisavalt, ribosoom seisma ei jää, jääb seisma alles
terminaator koodoni peal, see lõpetab valgu sünteesi. Moodustub alternatiivne
sekundaarstruktuur, terminatsiooni juuksenõel. Selle tekkimine sõltub sellest, kus ribosoom
jääb RNA peal seisma.
Transkriptsiooni terminatsiooni signaal on RNAs ja väljendub RNA ruumilise struktuuri tasemel.
Ribosoom muudab mRNA struktuuri – võimaldab või takistab terminaatori teket.
Erinevad viisid transkriptsiooni aktiveerimisel
: RNA polümeraasi seondumine erinevate
promootoritega sõltub sigma faktorist. Et suurendada transkriptsiooni initsiatsiooni tõenäosust
on võimalik suurendada RNA polümeraasi seondumise stabiilsust DNA peale
aktivaatorvalkudega. Nad seonduvad DNAga ette poole, kui RNA polümeraas ja on ühtlasi ka
seotud RNA polümeraasiga.
Translatsiooni repressioon valgu ja katalüütilise RNA poolt
: Trüptofaani operoni pole,
sünteesitakse mRNA, trüptofaani sünteesitakse palju. Trüptofaan seondub trüptofaani
repressorvalguga ja repressor promootorpiirkonnaga. Sünteesitakse mRNA, selle pealt GlcN6P,
see seondub mRNA kindla struktuuri elemendiga ja suunab mRNA struktuuri muutuseni nii, et
mRNAs tekitataks katkemine (ribolülitit), ta lagundatakse. RNA sünteesib RNA lagundamist, see
sõltub GlcN6P olemasolust.
Ribolülitid
: Nukleiinhappe struktuur sõltub temperatuurist, temperatuuri tõustes kaksikahela
paarid sulavad lahti ning sinna saavad seonduda ribosoomi subühikud, sest need ei saa
seonduda kaheahelalised RNAga. Kõrge temperatuuri puhul saab toimuda translatsioon.
Madalal temperatuuri on ribolülitid välja lülitatud. Kui tRNA otsas ei ole glütsiini, siis ta saab
paarduda, kui on, siis ei saa paarduda ja tekib terminatsiooni juuksenõel, terminatsioon, RNA
süntees jääb seisma. RNA struktuur sõltub mingi madalmolekulaarse ühendi sidumisest RNAle
või temperatuurist. Võib tekkida ka juuksenõel, mis peidab ribosoomide seondumise koha.
o Trüptofaani süntees lülitatakse välja ka repressorvalgu abil. Kui
trüptofaan sendub dimeersele trüptofaani repressorile(tase peab olema
piisavalt kõrge, siis see repressorvalk aktiveerituna seondub DNA-le ja
ei lase transkriptsioonil toimuda.
35
o Teistsugune regulatsioon toimub Glükoosamiin-6-fosfaadi puhul: siin on
ensüüm, mis teda aitab sünteesida, vastav süntaas.
Vastava geeni mittekodeerivas piirkonnas, kus veel see struktuurgeeni
pole algatatud, omandav see järjestus omamoodi struktuuri, kuna sinna
ribosoom ei seondu. Saab toimuda rahulikult glükoosamiini süntees
ning ta tase kõrgeneb rakus. Aga kui see tase on nüüd kõrge, siis
glükoosamiin seostub selle sama RNA-ga, millelt ta sünteesitakse. Nüüd
on RNA aktiveeritud nii käitub ennast hävitavalt, nimelt ta hakkab
ennast tõepoolest hävitama. Ning selliselt defektselt RNA-lt mingit
sünteesi ei toimu.
Neid nimetatakse ribolülititeks. Nad toimivad tänu RNA struktuuri
muutustele induktorite toimel:
o Üks ribolülitite näide oleks termosensori näide. On mikroorganismide
kaitse termilistele muutustele. Madala temp juures on struktuur üks,
kõrgemal teistsugune. RBS – ribosoomi seondumiskoht – RNA järjestus,
millega ribosoom seondub. Külma ilma struktuuris on struktuur
mittekättesaadav, kuid tõustes struktuur avaneb ja ribosoom seondub
RNA-ga ning saab toimuda vastav valgusüntees.
o Teine näide, T-box näide. tRNA sõltub struktuurielement: T-box on
mRNA osa. Kui selle glütsiinispetsiifilise tRNA hulk on rakus piisavalt
suur(Gly ple seal), siis seondub mRNA T-boxiga ka ning tekib
antiterminatsiooni struktuur.
Kui Gly tRNA on aminoatsüleeritud ehk tal on glütsiin otsas, siis mRNA-
ga ei ei ta seotud ning struktuur teistsugune ning terminaatorpiirkond
on olemas. Transkriptsioon on peatunud antud juhul.
LysC on ensüüm, mis on vajalik lüsiini biosünteesi ahelas.
S-adenosüülmetioniin SAM – väga tähtis kofaktor metüülrühmade ülekandel. Kui DNA
replikatsioonil või histoonidel modifikatsioonil on SAM just metionüülrühma doonor. SAM on
kindla kontsentratsiooniga rakus, et saaks nukleiinhappeid modifitseerida. Seda kontrollitakse
ribolüliti abil.
S-adenosüülhomotsüsteiin SAH – sama nagu SAM, aga metüülrühm ära
võetud.
Ribosoomide biosüntees
: Ribosoomide kontsentratsioon rakus võib muutuda väga suures
ulatuses. Ribosoom koosneb kahest subühikust. RNAst ja valkudest mõlemad subühikud. Igat
valku ja RNAd on ribosoomis üks koopia. Mõlemaid subühikuid peab olema võrdselt. Kui mõni
valk jääb sünteesimata, et ei saa kokku panna funktsionaalset ribosoomi.
Ribosoomi valkude süntees
: enamus ribosoomi valke on organiseeritud operonidesse. Nende
sünteesi regulatsioon seisneb tagasiside abil reguleerides. Produkt ise kontrollib enda sünteesi.
Enamus operone sisaldavad kas ainult ribosoomi valke. S10 operon kodeerib paljusid ribosoomi
valke. Üks neist – valk L4 – on võimeline kontrollima selle operoni avaldumist. Kui ta on
seondunud operoni regulaatorpiirkonnale, siis translatsiooni sealt ei toimu. Et sünteesida
operoni viimast valku, tuleb ikkagi alustada algusest ja sünteesida ka kõik eelnevad valgud,
sisemist translatsiooni initsiatsiooni ei ole.
36
Valgu S8 seondumine 16 S rRNA ja mRNaga
: väiksema subühiku valk, seondub 16S rRNA
mingi kindla piirkonnaga. Kui S8 on vähe rakus, siis ta seondub kõigepealt 16S rRNAga. Kui aga
teda on palju, siis ta seondub ka S8 mRNAga ning lülitab selle välja.
Pre-rRNA struktuur
: Ribsoomi RNA geenid on kõik ühes operonis ja sünteesitakse kõik ühe pika
eellasena. 2 promootorit, mis on tundlikud võlutäpikese suhtes. Kõige esimesena lülitatakse
rRNA süntees välja kui vaja. 16S ja 23S subühikud. RNaas III lõikab välja 16S ja 23S RNA
esialgsest pikast RNA transkriptist (lõikab katki kaheahelalised „raamjärjestused“). Lõplik
molekul moodustub RNaas E, RNaas G ja RNaas T toimel. rRNAle lisatakse modifitseeritud
nukleotiide vastavate ensüümide poolt ribosoomi kokkupaneku käigus – posttranslatsiooniline
modifikatsioonid (nt SAM, mis osaleb rRNA modifitseerimises).
23S rRNA protsessimine
: Seal on üks nukleotiid, mis ei ole paardunud, seda tunnebki ära
RNaas3.
Ribosoomide biosüntees
: rRNA süntees on rangelt kontrollitud: kasvukiirusest sõltuv
kontroll; „nälgimise korral lülitatakse välja (võlutäpikese kaudu); reguleeritakse vastavalt
translatsioonilisele aktiivsusele. Selles on väga palju erinevaid positiivse kui ka
negatiivse kontrolli elemente, mis aitavad ribosoomi sünteesil kiiresti välja lülituda.
Ribosoomi-valkude süntees järgib rRNA tootmist „ tagasiside“ kaudu. Kui rakkudse on
vaba rRNA siis toimub r-valkude sünteesi derepressioon. R-valkude negatiivne tagasiside
toimub nii translatsiooni kui transkriptsiooni tasemel.
Trankriptsioon eukarüootides
:
3 erinevat DNA sõltuvat RNA polümeraasi, bakterites on ainult 1.
Pol I transkribeerib vaid ühte geeni, mida on aga paju koopiaid, rRNA geene ainult
(rDNA).
Pol II transkribeerib enamikku geene ja toodab pre-mRNA. Enamus genoomist on RNA
polümeraas II poolt transkribeeritud. Ainult tema produktide pealt sünteesitakse valke.
Pol III transkribeerib 5S rRNA geene, tRNA geene ja snoRNA geene.
Geenid jagunevad eri liikideks produkti alusel
:
•
RNA geenid: rRNA geenid – Pol I;
tRNA, 5S rRNA, snoRNA geenid – Pol III;
miRNA ja teiste mittekodeerivate
RNAde
geenid (Pol II);
•
Valgugeenid: Pol II poolt transkribeeritavad.
•
„Koduhoidjate“ valgugeenide (poly(Py) trackt 5’ UTRis) intronites on sageli
väikeste RNAde geenid
Pseudogeenid
: geenid, mis ei kodeeri funktsionaalset produkti
(valgugeenidel enamasti raamis
stop koodon). Jagunevad protsessitud ja protsessimata pseudogeenideks, millest esimesed on
tekkinud RNA vahendusel. Suurem osa kordusjärjestustest koosnevad eri liiki pseudogeenidest.
Pseudogeenid võivad olla protsessitud(intronid eemaldatud) või protsessimata. Protsessitud on
tekkinud RNA vahendusel.
37
Pol II promootor
: Võrreldes bakterite promootoriga on RNA Pol II promootrid kompleksemad ja
sisaldavad konserveerunud järjestusi. Promootori erinevaid osi tunnevad ära erinevad valgud:
tähtsamad TFIIB ja TFIID. TBP(TATA binding proteiin) on valk, mis on seotud mitte ainult RNA
Pol IIga, vaid ka teiste RNA Polümeraasidega. TFIIB on kompleksne, koosneb väga paljudest
subühikutest (TAF – transkriptsiooni aktiveerivad faktorid).
Eukarüootide transkriptsiooni initsiatsioon
: palju rohkem lisafaktoreid kui bakteritel.
Transkriptsioonifaktorid seonduvad ka ilma RNA polümeraasita, bakteritel ei seondu. TBP ja
TFIID seonduvad DNAga ilma RNA polümeraasita, kõigepealt TFIID koos TBPga, siis TFIIA ja
TFIIB, siis alles RNA polümeraas koos TFIIFga. RNA polümeraas II hiiresaba
(karboksüterminaalne domään). Sellega seonduvad paljud erinevad valgud, TFIIE ja TFIIH (eriti
oluline), kui need seonduvad, moodustub avatud kompleks, aga transkriptsioon veel ei alga,
sest karboksüterminaalne domään on seotud üldiste transkriptsiooni faktoritega. Et
transkriptsioon algaks on vaja karboksüterminaalse domääni Seriini jäägid fosforüülida ja
seejärel karboksüterminaalne domään vabaneb üldiste transkriptsiooni faktorite kompleksist ja
algab transkriptsioon.
TBP on valk, TATA binding protein, seostub DNA TATA
järjestusega, kõverdab DNA ja see aitab kaasa
transkriptsiooni mulli tekkimisele. TBP on kõigi RNA
polümeraaside komponent, osaleb nii Pol I, Pol II kui Pol
III transkriptsiooni faktorina.| on valk, mis seondudes
DNA-ga põhjustab DNA kõverdumise, muudab ta kuju.
Kui on olemas TATA-boks, siis seostub sellega, kui seda
parajasti pole, siis kuhugi mujale. See kõverdumine on
vajalik avatud kompleksi tekkimiseks.
Kui me ahelad lahutame, siis ahelad pöörduvad kui nad
on lahti, kui otsad on aga kinni, seotud millegagi, siis
DNA ümber oma telje pöörelda ei saa ning läheb
krussi. Ahelate lahutamiskoha ees tekivad
positiivsed
supervindid, mis takistavad ahelate lahutamist, taga
tekivad
negatiivsed supervindid, mis soodustavad
ahelate lahutamist. Transkriptsioonilised
topoisomeraasid lasevad vindid välja, kui neid liiga
palju on. | Transkriptsiooni käigus muutub DNA
topoloogia. Kui DNA-d lahti harutada, siis
transkriptsioonimull muudab DNA struktuuri. Mulli ees
positiivsed supervindid, mulli taga tekivad aga
negatiised supervindid. Neid vinte tuleb kompenseerida, selleks on olemas topoisomeraas I ja
II.
Mediaator kompleks mõjutab transkriptsiooni initsiatsiooni. Üldised transkriptsiooni faktorid on
ühtemoodi olulised kõigi promootorite korral. Promootorist eemal, aga siiski samal DNA
molekulil asuvad
cis elemendid ja
trans faktorid(valgulised, lahustuvad transkriptsiooni
faktorid), mis seonduvad cis elementidega. Aktivaatorvalk seondub DNAga ühe otsaga ja
mediaatoriga teise otsaga. HAT histoonideatsetüül transferaas, et histoond laseksid lahti DNA
küljest nad atsetüleeritakse.
38
Mediaator kompleksid koosnevad paljudest erinevatest valkudest. Erinevates
organismirühmades on nad erinevad.
Faktorid, mis stimuleerivad transkriptsiooni elongatsiooni
: karboksüterminaalne domään on
fosforüleeritud transkriptsiooni alustamiseks. Ta on 7 AH koosnev korduvjärjestus. Kui
transkriptsioon jätkub, siis toimub karoksüterminaalse domääni defosforüülimine. Sellega aga
seonduvad paljud teised faktorid, mis on vajalikud RNAga toimuvateks sündmusteks.
Capping ensüüm – paneb Capi RNA Pol II produktidele otsa. Cap on mRNA 5’ otsas asuv
struktuur. Cap lisatakse kõigile RNA Pol II produktidele. RNA Pol II produktid algavad alati
puriiniga. Kohe kui RNA väljub RNA Polümeraasist, siis mRNA capping ensüüm liigub RNA külge
ning paneb Capi ta külge. Kui Capi pole, siis translatsiooni ja protsessimise(splaissimine,
polüadenüleerimine, RNA lõikamise ja spetsiifika faktorid) masinavärk seda RNAd ei puutu, nii
märgitakse ära, et tegu on RNA pol II produktiga. Kõik valgud, mis seostuvad pre-mRNAga
seonduvad kõigepealt karboksüterminaalse domääniga ning selle pealt lähevad üle pre-mRNA
külge.(eukarüootse mRNA otsas on (3’)m7GpppPu(5’) järjestus, mis pannakse sinna selle
ensüümi abil).
Eukarüootid trankriptsiooni terminatsioon
: RNA pol II transkribeerib RNAd, tema
karoksüterminaalse domääniga seostuvad valgud, mida kututakse CPSF. Väga tähtis. Põhiline
ensüüm, mis tunneb ära polüadenüleerimis signaali, määrab polüadenüleermiskoha ja lõikab
RNA katki. CstF
lõikamist stimuleeriv faktor. Nad on mõlemad karoksüterminaalse domääni
küljes kinni. Kui CPSF leiab sünteesitud RNA pealt polüadenüleerimise signaali, siis laseb RNA
polümeraasi küljest lahti ja seondub sünteesitava RNAga, sellega seondub ka CstF. CPSF lõikab
RNA lahti RNA polümeraasi küljest. RNA molekul on valmis. Terminatsiooni signaal tuntakse ära
RNA tasemel nagu bakteriteski.
CPSF suunab ka RNA pol II produkti polüadenüleermist, seda viib läbi poly-A polümeraas
(PAP)
, mis lisab RNA otsa polü A järjestuse (AAA), CPSFist sõltuv. Sünteesitava polü-Aga
seonduvad valgud, mis on polü-A seonduvad valgud, mis on olulised translatsiooniks,
splaissimiseks ja mRNA transpordiks (et tuumast transporditakse tsütoplasmasse).
RNA polümeraas pärast RNA ära lõikamist jätkab transkriptsiooni, võib transkribeerida kuni
paarsada nukleotiidi veel. Temaga aga ei ole enam seondunud faktorid, mis tagaksid RNA pol II
kõrget protsessiivsust. Kuna protessiivsus langeb, siis ta varem või hiljem lõpetab
transkriptsiooni ja nendel transkritidel pole olulist bioloogilist tähtsust. Sellel sünteesitaval jupil
pole ka Capi otsas, temaga ei seondu kaitsvaid faktoreid ning ta lagundatakse ära.
RNA Pol I promootor ja initsiatsioon
: sünteesib rRNAd. Promootorist 100-150 aluspaari eespool
on piirkond(UCE element), kuhu seonduvad üldised transkriptsiooni faktorid. Promootor ulatub
veidi transkriptsiooni saiti sisse. Pol I produktid moodustavad tuumakesed, milles toimub
ribosoomide süntees. Moodustuvad ribosoomide eellased. rRNA geenid paiknevad
tandemkordustena – promootor-struktuurgeenid|promootor-struktuurgeenid|jne. Iga
tandemkordus sisaldab endam 18S rRNA, 5.8rRNA, ja 28S rRNA-d. Nende vahel on spacer
järjestused. 5S rRNA-d transkribeerid hoopis pol III.
RNA Pol III promootor ja initsiatsioon tRNA geenidel
: produktid on väiksesed RNAd. Tal on
kahte erinevat tüüpi promootoreid sõltuvalt sellest, mida sünteesid. Tema produkte
modifitseeritakse teist moodi. Promootorid mõlemad asuvad transkripti sees(sisemine
promootor).Transkriptsiooni faktorite hulgas on TFIIIA (5SRNA) ja TFIIIC (tRNA jaoks).
39
Transkriptsiooni faktorid seonduvad
promootoriga ja ka produktiga. 5SRNA läheb
koos kompleksis TFIIIAga kompleksis
tuumast välja ja ribosoom pannakse kokku
alles tsütosoolis. Ribosoomi eellased
tehakse tuumakeses, kui ribosoomile
pannakse 5S valgud külge tsütosooli ja seal
saab ribosoom alles valmis.
Eukarüootide transkriptsiooni regulatsioon
:
regulatsioon toimub põhiliselt
transkriptsiooni initsiatsiooni reguleerimise
kaudu (preinitsiatsiooni kompleksi
moodustumine, suletud kompleks), kuid ka
transkriptsiooni elongatsiooni tasemel. Et
regulaatorvalgud leiaksid üles endale
vastavad cis elemendid tuleb DNA
kromatiidist välja õngitseda. | Inimesel üle
25000 valgu geeni. Ja 100 000 eri valku.
Vaja on seega alati sünteesida vajalik kogus
mRNA-d ja vajalikus koguses valke. Geene peab
aktiveerima eukarüootidel. Sest nad kompleksis
valkudega nukleosoom, histoonid. Peab lahti
pakkima, et neile ligi üldse saada. Suurel
määral kontroll käib nii, et otsustatakse kui
palju produkti on üldse tarvis sünteesida.
Transkriptsiooni regulatsioon toimub cis
elementide(promootor, terminaator jm.) ja
trans faktorite(valgud, mis DNA-ga seonduvad
või
ka kromatiiniga) interaktsiooni kaudu.
Kromatiini struktuur
Kogu DNA on pakitud nukleosoomidesse, mis
on omavahel tihedalt seotud 30 nm fibrilli, mis
moodustab omakorda järgmise taseme. 30 nm
fibrill on see, mis kujul on DNA tavalistes raku
tuumades.
1.
Selleks kõigepealt aktivaatorvalk seostub oma spetsiifilise DNAga DNA replikatsiooni
käigus.
2.
Peale replikatsiooni, selle koha peal, kus aktivaatorvalk seostus, tekib kromatiini
struktuuri muutus, seda katalüüsib CRC.
3.
Histoonide modifitseerimine, eelkõige H3 N-terminaalne saba modifitseeritakse, mis on
signaaliks järgmiste aktivaatorvalkude seondumisele.
4.
Tekib aktiveeritud geeni kompleks.
5.
Tekib transkriptsiooni pre-initsaitsiooni kompleks, mis sisaldab ka RNA polümeraasi
40
6.
Teised aktivaatorid seonduvad, karboksüterminaalse domääni fosforüülimine ja algab
transkriptsioon.
Interferool gamma geen on lähestikku toodud interlekeeni geenidega (suunavad
immuunvastust). Kui tekib aktiveeritud immuunvastus, interleukeeni geenid vaigistatakse.
Aktivaatorid on sarnase struktuuriga nagu bakteritel, koosnevad kahest domäänist. Üks
seostub DNAga ja teine on märklauaks RNA polümeraasile kas otse või mediaatori vahendusel.
Aktiivne vorm on aktivaatori dimeer.
Gal4
on spetsiifiline transkriptsiooni faktor. Üldised transkriptsiooni faktorid ei ole piisavad, et
GAL1 geeni peal transkriptsiooni alustada. Gal4 faktoritele on 4 sidumiskohta, iga sidumiskoht
seob dimeerse Gal4, kui neid on küllalt palju, siis hakkab kromatiini remodulleerimise kompleks
muutma DNA struktuuri ja muudab ta kättesadavaks üldistele transkriptsiooni faktoritele.
Modullaarsed valgud koosnevad kahest moodulist – DNAd siduv- ja aktiveeriv moodul.
Aktivaatorid koosnevad samuti moodulitest. Üks moodul on üks struktuurne domään. Moodulid
on omavahel seotud liikuva linkeriga (ühendav regioon). Aktivaatorid moodustavad dimeere,
korduvjärjestused. Mooduleid vahetatakse, kombineeritakse. Trankskriptsiooni aktivaator Gal4
seondub oma kindlale saidile. Kui võtta seal transkriptsiooni domään ja LexAst DNA sidumise
domään, siis moodustub uus aktivaator, kus lacZ geen on kontrollitud Gal4 aktiivsuse poolt.
Uus valk võtab omaks Gal4 omase signaali.
Kui meil on transkriptsiooni sait Gal4 ja sellel aktivaator: kui võtame kas seondumisdomääni või
aktivaatori domääni ja LexA saidi seondumisdomääni, siis saame kimäänse valgu, mis võtab
vastu Gal4 iseloomuliku transkriptsiooni aktivatsiooni signaali (mis on sisuliselt nälgimine).
Promootor (LexA) võtab seega vastu Gal4 signaali. |
Tranksriptsiooniaktivaatoritel 2 domääni: DNA-seondumisdomään ja
aktivaatorregioon. Need valgud aktivaatorid töötavad, kui modulaarsed
ühikud. Üks moodul ongi DNA-ga seonduv, teine on transkriptsiooni
reguleeriv moodul. Mooduleid saab kombineerida, ühendada eri mooduleid
ja eri moodulite kombinatsioonid viivad läbi erinevat, uut tüüpi
regulatsiooni.
Kui me nt Gal4 saidis on olemas vaid vastav DNA-sidumisdomään,
siis aktivatsiooni ei toimu, peab olema koos.
Meil võivad aga olla sellised valgud, mille aktivatsioonidomään võib
olla Gal4 oma, aga DNA-siduvas domäänis aga miski muu valk, nt
LexA, siis toimub siiski aktivatsioon. Nii saab ka uusi valke kodeerida
vajadusel.
Sellist aktivatsiooni võib läbi viia nt mediaatorkompleks.
Transkriptsiooni aktivatsioon võimendi abil
: TFIID seondub TATA piirkonnaga, aktivaatorid, mis
seonduvad võimendiga (transkriptsiooni cis element), mediaator seondub ka võimendiga, RNA
polümeraas TFIIDga. Geeni aktivatsioon saab toimuda siis, kui mediaator seob omavahel
võimendi ja promootori. Mediaatorkompleksi erinevad komponendid seonduvad erinevate
transkriptsiooni faktoritega ja viivad RNA polümeraasi aktiveerimisele.
Kromatiini remodeleermine ehk struktuuri muutus toimub selle käigus. Modifitseeritakse
histoone. Kui DNAga on seotud aktivaator valk, siis sellega seonduvad histoonide
41
atsetüültransferaasid ja modifitseerivad selles kromosoomi piirkonnas nukleosoomid. See on
signaaliks järgmistele valkudele, mis moduleerivad kromatiini edasi.
Pärmi HO geeni aktivatsioon: ta on endonukleaas, vajalik pärmirakkude jagunemisel. Seda
reguleerivad kaks transkriptsiooni aktivaatorit SWI5 ja SPF. Ho geen on vajalik, et kromosoomid
saaksid lahku minna. Ho endonukleaas avaldub ainult emarakkudes. Tütarrakus avaldub alles
siis, kui tütarrakk ise hakkab poolduma. Avaldub enne raku jagunemist. Kui SWI5 on
seondunud, siis toimub histoonide atsetüleerimine ja kromatiini moduleerimine. Seondub SPF,
mis seondub juba kromatiini lähedase piirkonnaga. Siis seondub juba mediaator kompleks ja
sellega RNA polümeraas.
Histoonide modifikatsioonide kaks toimet
•
DNA-Histoon seondumise nõrgenemine (nukleosoomi sees ja eriti nukleosoomide
vahelised interaktsioonid nõrgenevad)
•
Modifitseeritud histoonidega seonduvad mittehistoonsed valgud, mis
reguleerivad geeni avaldumist ja kromatiini remodelleerimise ensüümid
Histoonide modifikatsioonid
: H3 ja H4 moodustavad tetrameeri, mis jääb DNA replikatsioonil
DNAga seotuks. H2A ja H2B moodustavad kaks dimeeri, mis lahkuvad replikatsioonil DNA
küljest.
Eriti oluline on H3 modifikatsioon. Replikatsioon seda ei muuda. Histoonide globullarsest kehast
ulatuvad välja üheahelalised osad, N ja C terminaalsed otsad. Neid otsi modifitseeritakse,
lisatakse näiteks fosfaatrühm Seriinidele. Kõige olulisem modifikatsioon on atsetüleerimine ja
ka metüleerimine. On ka muid. Atsetüleermine ja metüleerimine toimub Lüsiini külgahela
aminorühmaga, pannake atsetüül otsa näiteks. Lüsiini lämmastik enam laengut ei oma ning
selline ilma laenguta osa ei saa DNAga enam seonduda. Kui lisatakse üks metüülrühm, siis
laeng jääb veel alles, kui aga lisatakse juba kaks, siis laeng kaob ning enam DNAga ei
moodusta ioonseid sidemeid. Metüleeritakse nii lüsiine kui arginiine, atsetüleeritakse ainult
lüsiine. Ubikvineeritakse ka lüsiine.
Histoonide atsetüültransferaas paned atsetüüli külge. Histooni deatsetülaas võtab
atsetüülrühma ära. Fosfaatrühmasid lisavad kinaasid, metüülrühmasid metülaasid. Histoonide
atsetülaasid on geenide regulatsioonis olulised valgud, sisaldavad ühesugust struktuurset
elementi – bromodomään.
Histooni H3 kovalentsed modifikatsioonid omavad erinevat tähendust
H3K9 histooni H3 9. lüsiin on kõige tähtsam geeni ekspressiooni seisukohalt. Histooni H3
kovalentsed modifikatsioonid omavad erinevat tähendust. Kui 9.lüsiin on atsetüleeritud, siis
42
nukleosoom muutub DNA suhtes liikuvaks, DNAd on võimalik vabastada nukelosoomide küljest.
On oluline transkriptsiooni elongatsiooni käigus
Kui histoone pole üldse modifitseeritud, siis sellises kromatiini piirkonnas geeni ekspressiooni ei
toimu, geenid on vaigistatud.
Kui H3 9.lüsiin on metüleeritud, siis ei saa teda ka enam atsetüleerida ning seda kromatiini
piirkonda ei saa aktiveerida, geenid on vaigistatud ja see kromatiiini piirkond moodustab
heterokromatiini.
Fosforüülimine toimub põhiliselt mitoosi käigus.
Modifikatsiooni saab lisada ja ära võtta, metüleermist on raske ära võtta, aga siiski võimalik.
Atsetüleerimist ja fosoforüleerimist saab ära võtta väga kergelt.
Lüsiinide ja arginiinide modifitseerimisel on kahesugune toime
. Esiteks mõjutavad histoonide
ja DNA vahelisi interaktsioone, atsetüleeritult ei moodusta ioonest interaktsiooni. On oluline
nukleosoomi enda ja temaga seotud DNA kõvasti sidumisel. Ioonsed interaktsioonid hoiavad
DNA keerde pinge all. Teine oluline asi on see, et erinevate nukleosoomide omavahelised
interaktsioonid, sest histooni saba ei seostu ainult DNAga, vaid ka kõrval oleva teise histooniga.
Regulaatorvalkude seondumise kohad: Regulaator valgud tunneb ära kindla modifikatsiooni
mustri. Oluline on, milline on histooni kõikide modifikatsioonide muster kokku. Erinevad
modifikatsioonid võimendavad näiteks üksteist. 9.lüsiini metüleerimine ja 10.seriini
fosforüülimine on negatiivse toimega näiteks ehk siis kui üks on toimunud neist, siis teist väga
kergesti enam kõrvale ei toimu.
Võimendi mõju tühistamine insulaatori abil: Geeni regulatsiooni cis element võimendi
võib olla promootorist kaugel kuni 50 000 aluspaari kaugemal, aga toimib ainult sama
molekuli piire. Võimendi aktiveerib mõlemas suunas geene DNA molekulil. Kui aga
promootori ja võimandi vahel on insulaator, siis see tühistab selles suunas võimendi
mõju ja promootorit enam ei aktiveerita. Insulaator on mingi kindel DNA järjestus, millega
seonduvad järjestuse spetsiifilised valgud.
Geeniklastreid reguleerib sageli LCR või GCR: imetajate globiini geenid. Imetajad saavad oma
verre hapniku ema organismist. Platsentas toimub hapniku ülekanne ema hemoglobiinilt loote
hemoglobiinile. Kuna lootesse on hapnikul pikk maa minna, siis on vaja, et hemoglobiin seoks
hapniku palju stabiilsemalt, kui iseseisvalt hingavas organismis. Sõltub hapniku ja CO2
partsiaalrõhkudest. St on vaja, et embrüonaalses staadiumis oleks teistsugune hemoglobiini
geen. Heem on ikka samasugune koguaeg. Imetajatel on vaja embrüonaalset hemoglobiini ja
täiskasvanule vajalikku hemoglobiini. Sünni momendil on vaja veel oma ülemineku
hemoglobiini, mis oleks vahepealsete omadustega. Kõikide globiini geenide lookust kontrollib
üks regioon LCR (lookust reguleeriv regioon). Hakkab avalduma see globiini geen, mille
läheduses on LCR. Embrüonaaalses staadiumis avaldub epsiloni geen, LCR on seotud epsilon
globiini geeniga. Edasise arengu käigus nihkub LCR seotuks gammaga, see on hilise
embrüonogeneesi ajal. Delta on sünni momendil. Beeta globiini geeniga on on LCR seotud
edaspidi ja jääbki sellega seotuks.
Hox geenide lookus sisaldab homeoboksi, määravad väga paljude teiste geenide aktiivsust.
Ülemregulaatorid. Aga mitte igasuguste geenide, vaid nende, mis on olulised keha plaanide
43
välja kujunemisel. Määrab milline komplekt geene, millises kohas avalduvad. Sarnane
regulatsioon LCRiga
Võimendi võib kombineeruda spetsiifiliste regulaatoritega: Võimendi läheduses on teiste
transkriptsiooni aktivaatorite seostumise piirkonnad. NFxB, IRF, ATF ja Jun on väga paljude
geenide transkriptsiooniga seotud.
Aktivaatorite kooperatiivne mõju: Kui kaks aktivaatorit seonduvad, kuid on üksteisest kaugel,
siis on vaja veel ühte faktorit, mis need kaks algset omavahel ühendaksid ja alles siis need
toimivad. Või kui üks aktivaator seondub, siis seondub kromatiini remoduleeriv kompleks ja siis
saavad alles teised aktivaatorid seonduda.
Repressorid võivad represseerida mediaatorid, polümeraasi, jms, pm kõike millega on võimalik
aktiveerida.
Eukarüootide pre-mRNA splaissimine - kokkulõikus
Peale RNA valmissünteesimist enamust RNAst protsessitakse, lõigatakse pikast eellasest välja.
Splaissimine – intronite eemaldamine, kõige olulisem protsess. Intronid võetakse välja ja
eksonid ühendatakse, informatsiooni muudetakse. Intronid on mittekodeerivad osad.
Kõigepealt sünteesib RNA polümeraas primaarse transkripti täpselt üks ühele ja siis tehakse
RNA molekul ümber.
Introneid esineb kõikidel organismidel. Intronite otsmised nukleotiidid on sarnased kõigil. Nad
erinevad intronite välja lõikamise protsessi poolest erinevatel organismidel. Taime geeni looma
viia ei õnnestu, sest introneid ei lõigata välja. Loomast looma saab küll viia. Taimedel puudub
introneid välja lõikav järjestus –
hargnemiskoha järjestus.
Hulkraksetes on kõvasti rohkem introneid kui ainuraksetes. Pagaripärmi genoomis on 200
intronit, need on seotud päriliku informatsiooni säilitamise ja ülekandmisega. Intronid on
enamasti pikemad (1500 inimesel), kui eksonid (umbes 10 korda lühemad).
Eksonid ei ole enamasti seotud valgu ruumiliste struktuurühikutega, aga paljude intronite
olemasolu annab meile võimaluse erinevateks alternatiivsetes splaissimisteks.
Intron-ekson skeem
Geen algab alati eksoniga, siis intronid ja eksonid vahelduvad. Intronit ei ole võimalik välja
lõigata, kui tal on ainult üks ots, siis teda ei kutsuta ka introniks, seega peab ta kindlasti olema
kahe eksoni vahel. Introni välja lõikamise informatsioon asub intronis endas. Introni 5’ ja 3’
otsas ja 3’ otsa läheduses paiknev ala sisaldab väljalõikamise infot.
5’-ekson,intron,ekson,intron,ekson,intron,ekson-3’. Geen algab ja lõppeb
eksoniga!
RNA splaissimine
RNA splaissimine toimub erinevate mehhanismide abil. Meie jaoks olulisim toimub
splaissosoom
i abil eukarüootsete rakkude tuumas pre-mRNA splaissimine. On ka neid
introneid, mis lõikavad end ise välja. Isesplaissuvad intronid jagunevad kahte gruppi. Grupp
kaks ei vaja mingeid kofaktoreid, sünteesitud RNA katalüüsib ise oma intronite välja lõikamist.
Grupp üks intronid kasutavad lisaks G-nukleotiidi. Info introni välja lõikamiseks ja eksonite
44
ühendamiseks paikneb intronis. Kui intron satub teise geeni, siis ta lõikab ennast ise ka sealt
geenist välja. Mõned intronid rändavadki ringi ühest geenist teise. Isesplassuvad intronid
võivad sisaldada valgu geene, mis aitavad tal liikuda ühest kohast teise.
Grupp 1 isesplaissuvad intronid: leiti 1980 T. Cech’i laboris, kes uuris Tetrahymena
thermophila rRNA introni splaissimist. Ta võttis rRNA geeni, transkribeeris in vitro selle pealt
RNA ja vaatas, mis juhtub selle RNAga. Ta vajab välja lõikamiseks G-nukleotiidi või G-
nukleosiid.| G1 intronid võtavad spets sekundaarse struktuuri, kus introni algus ja lõpp tuuakse
kokku. Ta toob lähedale G-nukleotiidi sidumistasku ja suunava järjestuse. Siin 5’ otsa lisatakse
G-nukleotiid(sinna vahele). Eksoni 3’ OH atakeerib teise eksoni 3’ fosfaati. Introni 3’ otsas on
samuti G-nukleotiid, mis läheb taas sidumistaskusse, ekson lõikatakse seejuures ära. Intron ise
võtab tsirkulaarse struktuuri.
Ribosüüm – RNA põhine ensüüm.
Kõigil grupp 1 intronitele on sarnane ruumiline struktuur. Koosnevad kindlatest heeliksitest.
RNA ahelad paarduvad iseendaga, vahelduvad paardunud ja üksikahelalised osad. Kokku
moodustub globulaarne RNA molekul. RNA põhised ensüümid on globulaarsed. Sisemine giid
järjestus – määrab 5’ eksoni otsa, määrab ära kus tehakse esimene katkestus. G-nukleotiidi
sidumise tasku – oluline 3’ hapnik, seotakse G-nukleotiid (astub ise keemilisse reaktsiooni, st 3’
hapnik atakeerib introni 5’ otsa fosfaati). See tasku moodustub GC paari abil. Introni
tsirkulariseerimine on pöörduv, st et intron võib end avada ja selle tulemusena tekib inaktiivne
intron (ei saa midagi enam katalüüsida).
RNA katalüütiline aktiivsus põhineb RNA ruumilisel struktuuril.
Eukarüootide tuumas splaissivad intronid
: Kahte erinevat tüüpi. Tuntaks otsmiste järjestuste
järgi või mažoorsed ja minoorsed. Minoorsed on vähe esinevad.
Mažoorsed intronid(G2?) algab
GU…AG-ga lõpeb. Lisaks G nukleotiidile on väga oluline ka A
nukleotiid. Tal on hargnemiskoha järjestus, kus on A nukleotiidi 2’ hapnik atakeerib 5’ splaiss
saiti, tekib tsükliline RNA. 5’ eksoni 3’ ots atakeerib splaiss saiti, seetõttu kutsutakse splaiss
saite doonor-splaiss saidiks ja aktseptor-splaiss saidiks. Ümber esterfitseerimise tulemusena
eksonid ühendatakse ja introni vabaneb. Seda protsessi katalüüsivad põhilised RNA molekulid,
mis on kompleksis valkudega.
Splaissosoomi poolt katalüüsitav introni eemaldamine(sarnasus G2 intronitele):
hargnemiskoha A 2’ hapnik satub splaiss saidi lähedusse. Splaissimis koha A-l on nii 2’ kui ka 3’
hapnik ühendatud fosfaatidega. Splaissimise tulemusena võib ühe geeni pealt tulla palju
erinevaid valke.
RNA polümeraas II produkte splaissitakse. Splaissimist katalüüsivad 5 erinevat RNA molekuli ja
150 valku. Mõned 150st on seotud karboksüterminaalse domääniga, kui RNA valgud hüppavad
RNA polümeraasilt RNA peale, hakkab splaissimine. RNP ehk ribonukleotiidsed partiklid. U-RNAd
(1,2,4,5,6) osalevad otste trimmimisel. Esimene RNA, mis leiti, mida kututakse U1, kõikide
rakkude tuumades ühesugune, sisaldab ebatavaliselt palju U-nukleotiide üle 30 %. Tavalised
alla 25%. Nüüd kõik väiksesed tuuma RNAd on U-RNAd.
Osad intronite nukleotiidid on alati kõigil intronitel samad, kaks esimest, kaks viimast ja
hargnemiskoha A. Kuus esimest nukleotiidi on ka peaaegu alati samad. Polüpürimidiin järjestus
45
3’ otsas, sisaldab palju C ja U. Splaissimine algab, U1 RNA kompleksis valkudega moodustab
U1snRNP, mis seondub 5’ splaiss saidiga, mille kõigepealt tunneb ära U1(RNA-RNA
interaktsioon). Seondub veel valk U2aktiveeriv faktor, mis seondub polüpürimidiin järjestusega
(valk-RNA interaktsioon). Hargnemiskohaga seondub BBP valk(hargnemiskoha järjestuse valk).
Kõik need moodustavad kokku varajase kompleksi ehk A-kompleksi. Edasi BBp asendub U2
RNAga, mis paardub hargnemiskoha järjestusega, mis on universaalselt konserveerunud,
hargnemiskoha A ei ole paardunud, ta on välja lükatud, tema 2’ hapnik satub välja heeliksi
pinnale. Edasi seondub kompleks, mis sisaldab U4, U5, U6 RNAd , tekib B-kompleks. Nüüd
moodustub introni iseloomulik ruumiline struktuur. 5’ splaiss sait ja hargnemiskoha A satuvad
lähestikku. Edasi U1 RNA lahkub ja 5’ introni %’ otsaga jääb seotuks U6 RNA ja lahkub ka U4
RNA. U5, U2 ja U6 RNA jäävad introni külge. U2 ja U6 peale kokku moodustub katalüütiline
tsenter, mis katalüüsib esterfikatsiooni. Intorni 5’ ots atakeerib 3’ splaiss saiti, eksonid
ühendatake, intron lahkub lasso kujul.
SR valgud on eksoonsed võimendid. SR seriini ja arginiini rikkad valgud. Seonduvad mitte päris
splaiss saitidega, vaid nende lähedusse ja suunavad nende splaiss saitide eelsitatud
kasutamist, mille lähedal nad on.
ATAC intronid ehk minoorsed intronid
DNA tasemel esimesed nukleotiidid AT ja viimased AC. Väga sarnane mažoorsele välja
lõikamisele. Intronite äratundmise signaalid on hoopis teistsugused. ATAC intronite 5’ otsa ära
tundmiseks on U11 RNA, hargnemiskoha ära tundmiseks on U12 RNA. Aga valgulised
komponendid on suurel osal samad, RNA komponendid on erinevad. Kõigepealt tuntakse ära 5’
splaiss sait, siis hargnemiskoht , siis RNA struktuuri ümber korraldamine, 5’ ots tuuakse
hargnemiskoha juurde. Toimub lasso moodustamine. 2’ hapnik atakeerib 5’ fosfaati. Hapnik
atakeerib ka teist splaiss saiti. Väga sarnane mažoorsega.
Minoorsed introneid on inimesel paar % kõigist intronitest, nad on seotud info ülekandega
geenides. Nende välja lõikamise kaudu reguleeritakse mitmete geenide avaldumise taset. Võib
olla ka geene, millel on 1-2 minoorset ja ülejäänud intronid mažoorsed.
Metalliioonidel väga tähtis osa RNA katalüüsimisel. Ribosüümid katalüüsivad reaktsioone
kahevalentse metalliiooni juuresolekul. Splaissimisel osalevad metalliioonid, seonduvad
koordinatiivsete sidemetega introni kindlate piirkondadega. Nt Mg2+ ioone. Need metalliioonid
aitavad katalüüsi keemilist etappi läbi viia. Hargnemiskoht A peab deprotoneeruma ning selles
osalevadki 2-valentsed metalliioonid.
Post-trankriptsiooniline geeniregulatsioon eukarüootides
.
Kõigil organismidel on geene umbes sama palju. Inimesel on geeni produkte väga palju rohkem
kui ussikesel. Võib olla juba million erinevat immuunoglobuliini molekuli, kuigi geenide arv on
enam-vähem sama neil.
Alternatiivne splaissimine(isemoodi kokkulõikus)
: geen algab esimese eksoni algusest ja
lõppeb viimase eksoni lõpuga. RNA primaarne produkt on identne ühega kahest DNA ahelast.
Sellest primaarsest RNAst on võimalik saada palju erinevaid produkte. Näiteks mõlemad
intronid lõigatakse välja ja kõik kolm eksonit pannakse omavahel kokku. Või jäetakse näiteks
teine ekson välja ja ühendatakse esimene ja kolmas. 3’ splaiss saidi määramine pole nii täpne
kui 5’ oma, seega on palju krüptilisi splaiss saite ja võidakse kasutada erinevaid splaiss saite.
46
Seega võib juhtuda, et teine ekson on pikendatud esimese introni arvelt, produkt on jälle
erinev, sest osa intronist on alles. Võib ka nii, et kogu intron jääb alles vahele. Võib ka nii, et
tekib kaks produkti, kus esimene ekson on ühendatud teisega ja teises produktis esimene
ühendatud kolmandaga. Need on põhilised võimalused alternatiivsel splaissimisel.
Geeniregulatsiooni elemendid. Geenide avaldumise valvamine: Ka splaissimise regulatsiooni
puhul on cis elemendid ja trans faktorid. Trans faktorid on enamasti valgud, mis seonduvad
järjestuse spetsiifiliselt reguleeritava geeniga DNA või RNa tasemel. | Cis-elemendid asuvad
reguleeritava molekuli peal. Trans-faktorid on enamasti valgud ka vahel RNA molekulid. Nad
seonduvad Cis elementidega ning mõjutavad geeni avaldumist.
Soo määramise geenid
: Alternatiivne splaissimine toimub ka soomääramise geenidel.
Äädikakärbsel määrab soo geeni hulk. Sõltub geenide suhtest. Kui X:A=0,5, siis isane. Kui see
=1, siis emane.
Sex lethal geenis toimub isastel ja emaste erinev splaissimine. Produkt ei ole enam
funktsionaalne isaste puhul. Ja nii ka järgmise geeni puhul. Kolmanda geeni puhul võimendit
pole ja splaiss saiti ei aktiveerita nagu emastel seda tehakse.
Kui aga ühte splaiss saiti ei kasutata, tekib repressor valk ja represseerib järgmise geeni RNA
alternatiivset splaissimist, see on emastel. Represseeritakse esimese splaiss saidi 3’ splaiss
saiti. Tekib produkt, mis omakorda on aktivaator kolmandale geenile ja aktiveerib selle peal
splaiss saiti.
Tekib valk, mille esimesed 400 AH on identsed nii emasel kui isasel. Isasel lisaks 150 AHd.
Emasel ainult 30 AHd. See valk represseerib vastavalt siis kas isas või emas geenide
avaldumist. Geeni doos on erinev ja sõltuvalt sellest siis tekib kas emane või isane
Alternatiivse splaissimise positiivne ja negatiivne kontroll
: 3’ splaiss saidi juures toimub
enamus reguleerimisest. Kui 3’ splaiss saidiga seondub repressor, siis U2 ei saa sinna seonduda
ja splaissimist ei toimu. Aktivaatori seondumisel splaissimine toimub, ilma ei toimu.
Fibronektiini geen
: fibronektiin on valk, mis koosneb väga paljudest korduvjärjestustest.
Geenis on neid korduvjärjestusi eriti palju. Fibronektiin on vere valk, ta on fibroblastides
ekspresseeruv valk, seotud vere hüübimisega. Avaldub fibroblastides ja hepatotsüütides.
Erinevates kudedes on tal erinev järjestus. Maksa rakkudes on tal struktuurne funktsioon.
Fibroblastides peab ta aga hüübima. See saavutatakse splaiss saidi represseerimise teel, ühes
on välja lõigatud, teistes pole.
Puuviljakärbsel väga paljudest eksonitest valitakse välja ainult üks, enamus lõigatakse välja,
mis on kordused. See mis kus kasutusele läheb sõltub konkreetsetest oludest.
Alternatiivne splaissimine sisekõrva harjarakkude slo mRNA puhul
: Tigu on õige olulisem,
temas on harjarakud, mille iga raku otsas on väikene harjas või jätke, mis on erineva
pikkusega. Kui jätke on pikk, siis tema oma võnkesagedus on madal, kui lühike siis
võnkesagedus on kõrge. Kui harjaraku harja võnkumine läheb resonantsi väljast tuleva
võnkumisega, siis annab harja rakk edasi signaali kuulmisnärvile. Kõik harjarakud pärinevad
ühest eellasest. slo mRNA splaissimine toimub erineatel harjarakkudel erinevalt. | Kõrva
tundlikkuse suurendamiseks on karvad pidevalt kergelt võnkuvad. Ses mõttes teeb kõrv ise
häältki. Mõnel on see hääl niivõrd võimas, et tõsiselt häiriv. Ilmselt on see ka üheks
47
hullumeelsuse tekitajaks. Karvarakkudes avaldub geen slo. See geen on suurem osa raku sees,
aga väike osa ka rakust väljas. Sel geenil väga palju alternatiivse splaissimise variante. On
seotud karvapikkusega. Splaissimise saite on mitmeid. 17 000 eri saiti.
Alternatiivse splaiss saidi valik
:
Olulised valgud, mis määravad millist konkreetset splaiss-saiti alternatiivsete hulgast
kasutatakse on eksoonsed võimendajad. Eksoonsed võimendajad seonduvad eksoneis
asuvate RNA järjestustega.
Splaissimine tagab eri valkude sünteesi sama geeni pealt.
Kotranskriptsioonilised sündmused pre-mRNA sünteesil
Kõigepealt 5’ otsa lisatakse cap. Kõik kotranslatsioonilised sündmused on seotud RNA
polümeraas II ja tema karboksüterminaalse domääniga. Edasi toimub splaissimine ja
selle reguleerimine (seonduvad splaissimisfaktorid ja võimendid/repressorid, millest
mõnede seostumine käib üle RNA pol II karboksüterimaalse domääni). 3’ protsessimine ja
seejärel seonduvad järgmised elemendid, mis splaissitud RNA transpordivad läbi
tuumapoori tsütoplasmasse. Transleerimine.
Trans-splaissimine
: Esineb väga sageli eukarüootidel. Väga paljudel geenidel on valgu
mRNA esimene ots ühesugune. Geenid, mis asuvad genoomis väikeste vahedega samas
suunas, samal ahelal on ühine esimene ekson. Esimese eksoni järjestus on kodeeritud
eraldi geeni poolt, mis asub unikaalses lookuses. Trans-splaissimises ühendatakse kahe
erineva RNA järjestused. See esineb algloomadel, nematoodidel, lameussidel, imetajate
viirustel.
Varbussi näitel(C.elegans). 70% mRNA-dest kannavad 5’ otsas ühte kahesti 22nt
järjestusest(SL1>50% ja ülejäänud on SL2). Need liidetakse geeniproduktide 5’ otsa.
RNA toimetamine (editing)
: RNA järjestuse muutmine peale transkriptsiooni. Inimesel kahte
tüüpi: adeniini muutmine inosiiniks ja tsütosiini muutmine uridiiniks. DNA ja RNA ei ole enam
samasugused, pärilik info muutub. 50 geenis inimesel muutub adeniin inosiiniks. : adeniin
reageerib veega, deamineerimmine, aminorühm asendub hapnikuga.
mRNA translatsioon – muudab AH spetsiifikat, tekib terminatsiooni koodon. Terminatsiooni
koodon ei sisalda kunagi tsütosiini.
RNA struktuuri muutus – mõjutab valkude seondumist RNAga, näiteks splaissimisel olulised
valgud.
pre-mRNA splaissimise mustri muutus – alternatiivsete splaiss saidi kasutamine
RNA degradatsioon – kui adenosiini 6C aminorühm küljes on metüülrühm, siis suunatakse RNA
degratatsiooni
RNA replikatsioon – viiruste puhul
U nukleotiidi lisamine RNA tasemel
: U nukleotiidide lisamine ja ulatuslik RNA toimetamine
inimesel eriti ei esine, aga esineb näiteks taimede mitokondrites, ainuraksetel parasiitsetel
algloomadel (nt trüpanosoomid). Küpsesse RNAsse on lisatud U nukleotiide ühte kindlasse
48
järjestikusse piirkonda. U nukleotiidi lisamise tõttu kodeerib mRNA nüüd hoopis teistsuguseid
AHsid. Toimetamiseks on mitu signaali. Seda viivad läbi giid RNA molekulid, mis sisaldavad
ankur järjestust, millele järgneb toimetatav regioon. Ankurjärjestus paardub mRNA mingi
piirkonnaga ja giid järjestuse alusel tehakse mRNA järjestus ümber. Toimetamine annab
parasiitidele võimaluse antigeeni järjestusi muuta enda kasuks kiiresti, nad on
immuunsüsteemile väga rasked ära tunda, sest nad muudavad antikehade järjestusi.
Adenosiini deaminaas:RNA järjestuse muutmine RNA spetsiifiliselt toimub ensüümi adenosiini
deaminaasi abil. Deamineerimine toimub adenosiini reaktsioonil veega. Ainult kindlatel RNAdel
muudetakse järjestusi. ADAR1 ja ADAR2. Nad on sõltuvad kaheahelalisest RNAst. ADAR
muudab mingis kindlas piirkonnas adeniinid inosiinideks. See on signaalik RNA
degradatsioonile.
mRNA (cargo) transport ekportiini abil
: mRNA sünteesitakse tuumas, läbib splaissimise ja
mRNAga jäävad seotuks paljud valgud. Splaiss saidi ümbrusesse jäävad seotuks valgud. mRNA
5’ otsa CAPiga seondub eIF-4E. Ekportiinid vastutavad mRNA transpordi eest. 5 erinevat
eksportiini. Transpordivad mRNAd tuumast tsütoplasmasse ja lähevad ise tuuma tagasi. Valgu
lokalisatsioon on määratud tema järjestusega. Järjestus peab aga olema kätte saadav. Kui
eksportiinid on kompleksis RNAga, siis nende tuuma lokalisatsiooni signaal on peidetud ning
nad suunatakse tuumast välja. Kui nad aga RNA küljes vabanevad on nende tuuma
lokalisatsiooni järjestus vaba ning nad suunatakse tuuma tagasi. Tuuma lokalisatsiooni signaal
sisaldab 4-5 järjestikku asetsevad Arg või Lys (positiivselt laetud), mis RNA keskel asetsevad.
Valgu biosüntees
Nukleiinhappe nukleotiidne järjestus tõlgitakse valkude aminohappeliseks järjestuseks
geneetilise koodi alusel. Geneetiline kood on universaalne kõikide organismi rühmades. Aga on
ka erandeid.
Universaalne geneetiline kood
49
1.\
2.
tä
ht
U C
A
G 3.t
äh
t
U UUU
-Phe
UUC
-Phe
UUA
-Leu
UUG
-Leu
UCU
-Ser
UCC
UCA
UCG
UAU-
Tyr
UAC-
Tyr
UAA-
Stop
UAG-
Stop
UGU
-Cys
UGC
-Cys
UGA
-Stop
UGG
-Trp
U
C
A
G
C CUU
-Leu
CUC
CUA
CUG
CCU
-Pro
CCC
CCA
CCG
CAU-
His
CAC-
His
CAA-
Gln
CAG-
Gln
CGU
-Arg
CGC
CGA
CGG
U
C
A
G
A AUU
-Ile
AUC
-Ile
AUA
-Ile
AUG
-Met
ACU
-Thr
ACC
ACA
ACG
AAU-
Asn
AAC-
Asn
AAA-
Lys
AAG-
Lys
AGU
-Ser
AGC
-Ser
AGA
-Arg
AGG
-Arg
U
C
A
G
G GUU
-Val
GUC
GUA
GUG
GCU
-Ala
GCC
GCA
GC
G
GAU-
Asp
GAC-
Asp
GAA-
Glu
GAG-
Glu
GGU
-Gly
GGC
GGA
GG
G
U
C
A
G
1960ndate algusel tehti kindlaks geneetiline kood, kasutati selleks ribosoome ja täis või
poolsünteetilisi nukleiinhappe järjestusi, esimesena tehti kindlaks UUU koodon (vaadati,
millise AH sidumisel ribosoomidel stimuleerib polüU lisamisel), sellega tavaliselt
alustatakse tabelit seetõttu. Vaadati millise AH sidumist ribosoomile stimuleerib 3-
uridüülhape. Selles koodis on terve hulk seaduspärasusi. Universaalne, sest
prokarüootidel, eukarüootidel ja arhedel vastab kood samale aminohappele.
Organiseeritud kolme kaupa.
Geneetiliselt kodeeritud AHsid on kokku 22, aga kõik nad tavaliselt ei esine ühes organismis. 20
on tavaliselt kõik olemas suuremates valkudes, väikestes ei pruugi kõiki olla. AHde evolutsioon
on seotud geneetilise koodi evolutsiooniga. • geneetiline kood on universaalne – see viitab
koodi ja järelikult valgusünteesi aparaadi monofüleetilisele päritolule.
Kombinatoorika: On 4 nukleotiidi. 4^3=64 koodonite arv. 61 vastavad aminohapetele, 3 on
stopp koodonid UAA, UAG, UGA. Ühele AHle vastab rohkem kui üks koodon. Trüptofaanile ja
Metioniinile vastab ainult üks koodon. Seriini erinevad koodonid asuvad üksteisest kaugel,
ülejäänute koodonid asuvad lähestikku.
Kui koodonid on sarnased, mis kodeerivad samu AHsid, siis väikse vea tekkides jääb AH ikkagi
samaks. tRNA toob AH ribosoomi, et koodon on nii organiseeritud on võimalik kasutada vähem
erinevaid tRNAsid.
Sünonüümsed koodonid – samat AHd kodeerivad.
Erinevad AHde külgahelad on erinevate füüsikalis-keemiliste omadustega. Asn ja Gln on
sarnaste omadustega, asuvad koodi tabelis lähestikku.
Sagedastele AHdele Leu, Ser, Ala, Gly, Arg vastab palju koodoneid.
„Külmunud õnnetuse“ hüpotees – kui koodon kunagi tekkis, ta pidi fikseeruma, sest kui muutus
geneetilises koodis tooks kaasa muutuse kõikides valkudes, kus see koodon on sees. Aga
tegelikult on koodon ikkagi muutunud. Nt mitokondritel on enamasti erinev geneetiline kood kui
peremees geneetiline kood, va taimedel. See saab muutuda teistsuguseks, sest mitokondritel
on väga vähe valgu geene, ainult kuskil 20. Sellise väikese arvu geenide puhul saab tekkida
juhus, kus teatud koodon ei esine mitte üheski valgus ja kui ta esineb ainult ühes valgus, siis
see muutus ei too kaasa suurt õnnetust.
Selgroogsete mitokondri koodis on 4 stopp koodonit. Ühtegi AHd ei ole kodeeritud ainult ühe
koodoniga. Koodon on edasi evolutsioneerunud, muutunud lihtsamaks. Ei ole ühe koodoniga
AHsid, lihtsamini ära tuntavad kui rohkem ühesuguseid koodoneid. Mitokondrite translatsiooni
on võimalik läbi viia vähema arvu tRNAdega.
Selgroogsete mito kood
:
50
Koodonite lugemise reeglid
:
1.
Koodon-antikoodon äratundmisel toimub koodoni 1. ja 2. positsioonis Watson-Crick
tüüpi aluspaardumine. 3. positsioonis on lisaks GU või UG paardumine nn „wobble“ paar, 3.
positsioonis tekivad kas AG, GU, CA, või AC paarid. Reeglite taga on ribosoomis toimuv. Ta
reguleerib, kuidas koodon ja antikoodon paarduvad. Ribosoom kontrollib ja reguleerib täpselt
ka aluspaaride geomeetriat. Wobble saab toimuda tänu ribosoomi mitte nii täpsele(vaatab
aknast välja) kontrollile.
Koodon-antikoodon paardumine toimub antiparalleelselt.
Koodon-antikoodn seondumine tuntakse ära väikese vao poolt. A nukleotiid kontrollib mRNA ja
tRNA 2’hapnike vahelist kaugust. Kui see kaugus on liiga suur, siis ei saa nad paarduda.
Koodoni kolmandas positsioonis moodustatakse vesiniksidemeid ainult 3 U-ga.
Lugemisraam ORF (avatud lugemisraam)
Alustame kas esimesest, teisest või kolmandast nukleotiidist. Meil on 3 lugemisraami O, +1 ja -
1 raam. Kõigis raamides tulev valgujärjestus on täiesti erinev. Väga tähtis on, et lugemisraami
lugemine algaks õigest nukleotiidist ning translatsiooni käigus ei nihkuks paigast ära, säiliks.
Supressor tRNA - vajalik, et transleerida nukleiinhappe järjestus valgujärjestuseks nii, et see ei
vastaks universaalsele koodile. Nonsenss supressor tRNAd suudavad AHd lülitada stopp-
koodoni kohal, neil on muteerunud antikoodon, mis võimaldab koodoni ja antikoodoni Watson-
Crick paardumist esimeses ja teises positsioonis ja seega võimaldab see stoppkoodoni koha
peal AH lülitamist. Stopp koodoneid kutsuti ka non-sense koodoniteks vanasti. Supressorite
antikoodonis on toimunud mutatsioon .
51
aminohap
e A.K.
(koodon)
aminohap
e A.K.
(koodon)
aminohap
e A.K.
(koodon)
Aminoha
pe A.K.
(koodon)
Phe
(UUU)
Phe
(UUC)
GAA
Ser
(UCU)
Ser
(UCC)
Tyr
(UAU)
Tyr
(UAC)
GUA
Cys
(UGU)
Cys
(UGC)
GCA
Leu
(UUA)
UAA
Leu
(UUG)
Ser
(UCA)
UGA
Ser
(UCG)
Stop
(UAA)
Stop
(UAG)
Trp
(UGA)
UCA
Trp
(UGG)
Leu
(CUU)
Leu
(CUC)
Pro
(CCU)
Pro
(CCC)
His
(CAU)
His
(CAC)
GUG
Arg
(CGU)
Arg
(CGC)
Leu
(CUA)
UAG
Leu
(CUG)
Pro
(CCA)
UGG
Pro
(CCG)
Gln
(CAA)
UUG
Gln
(CAG)
Arg
(CGA)
UCG
Arg
(CGG)
Ile (AUU)
Ile (AUC)
GAU
Thr
(ACU)
Thr
(ACC)
Asn
(AAU)
Asn
(AAC)
GUU
Ser
(AGU)
Ser
(AGC)
GCU
Met
(AUA)
UAU
Met
(AUG)
Thr
(ACA)
UGU
Thr
(ACG)
Lys
(AAA)
UUU
Lys
(AAG)
Stop
(AGA)
Stop
(AGG)
Val
(GUU)
Val
(GUC)
Ala
(GCU)
Ala
(GCC)
Asp
(GAU)
Asp
(GAC)
GUC
Gly
(GGU)
Gly
(GGC)
Val
(GUA)
UAC
Val
(GUG)
Ala
(GCA)
UGC
Ala
(GCG)
Glu
(GAA)
UUC
Glu
(GAG)
Gly
(GGA)
UCC
Gly
(GGG)
Supressor tRNA esineb rakus lisaks tavalisele tRNAle. Ta on vähem võimekas, kui tavaline
tRNA. Supressor tRNA ei ole nii efektiivne, efektiivusus jääb alati alla 50%, st vahest lülitab AH
valgu koosseisu, vahest mitte, valgu süntees saab jätkuda. Kui valgu geenis on raamis stopp
koodon, siis valgu ekspressiooni tase küll lagenb, kui supressori olemasolu korral teda ikkagi
transleeritakse, välja ei lülitata täielikult.
Sec ehk selenotsüsteiin – üks kahest vähem levinud AHst. Kõikides selgroogsetes vajalik.
Esineb väheses valkudes, kuskil paarikümnes valgus esineb. Bakterid suudavad aeroobsetes
tingimustes elada ilma selleta, anaeroobsetes aga mitte. Bakteril 4 geeni, kus on Sec sees.
Sec lülitamiseks valgu ahelasse on vaja 3 geeni produkti:
1.
tRNA, mille antikoodon vastab UGA koodonile
2.
ensüüm, mis sünteesib Seci – sünteesitakse teda seriinist. Sec spetsiifiline tRNA
sünteesitakse, selle otsa pannakse seriin. SecS on see ensüüm.
3.
EF-Tu üks eriline vorm – valk, mis transpordib Seci tRNA ribosoomidesse ja aitab tarnal
seonduda ribosoomi A saiti.
4.
UGA koodoni alla voolu peab olema kindlal kaugusel juuksenõela struktuur, et saaks
Seci valgu ahelasse lülitada.
Päristuumsetel on see veel keerulisem .
Pürrolüsiin ehk Pyl on ka geneetiliselt kodeeritud AH. Esineb ainult arhedel. Lülitub valgu
ahelasse UAG stopp koodoni koha peal. Ainult mõnedel arhedel on Sec ja Pyl mõlemad.
Enamustes organismides on siiski 21 AHd.
Koodonite kallutatus
Koodonite kasutamise juures on mõjutegurid, mis suunavad koodonite kasutamist. Üheks neist
on genoomi suunatud mõjurid.
1.
Igal genoomil on oma iseloomulik
GC paaride protsent. Nt enamusel termofiilsetel
organismidel on väga kõrge GC paaride protsent võrreldes mesofiilsetega või krüofiilsetega.
Imetajatel on eelistatult AT paarid, GC paaride osa on suhteliselt väike. (G/C rikkuse määrab ära
mittekodeeriv osa genoomist, sest kodeeriva osa hulk genoomist on väike, 99% on
mittekodeeriv).
2.
Teiseks teguriks on
transkriptsiooni ja replikatsiooni mõju. Replikatsioonil on üks
ahel juhtiv ja teine „lohisev“ ahel. Juhtival ahelal asub rohkem kõrgelt avalduvaid geene. Kuna
kõrgel tasemel ekspresseeruvad geenid kasutavad vähem koodoneid, suunab see suurel
määral koodonite kasutamist. Lisaks on teatud järjestused välditud (ei esine), mis avaldab ka
mõju koodonite kasutamisele. Nt transkriptsiooni initsiatsiooni signaalid.
3.
Oluline nähtus on ka
horisontaalne geeniülekanne. Geenid rändavad ühest taksonist
teise. Kust geen pärit on peegeldub tema koodonite kasutamises.
CAI – koodonite adaptatsiooni indeks
Väärtus peegeldab geeni avaldumise tasemes. CAI on suur nendel geenidel, mis avalduvad
kõrgel tasemel ja madal nendel, mis avalduvad madalal tasemel.
Arvutamine: GCU, GCC, GCA, GCG (arginiini koodoni perekonnad), igale koodonile antakse
arvuline väärtus vastavalt sellele kui palju seda koodonit esineb antud geenis. Nt selles geenis
on 20 arginiini koodonit, kus GCUd on 10. Kui võtta sageduseks 0,5 siis GCU CAI väärtus on 1.
52
Iga koodoni indeksid korrutatakse ja saadakse korrutise tulemusena geeni adaptatsioon
indeksi.
CAI alusel saab ennustada geeni avaldumise taset. Kui väärtus on kõrge, siis avaldub geen
kõrgel tasemel.
Standardiseerimine
Nähtus, mis võimaldab rakkudel energiat kokku hoida. Kui kõrgelt ekspresseeruvad geenid
kasutavad väheseid tRNAsid, on rakkudes neid teatud tRNAsid palju ja seega on võimalik
translatsiooni optimiseerida niimoodi, et ei sünteesita kõiki tRNAsid ja translatsiooni kiirus on
kiirem.
Koodonite kontekst
Kasutatakse eelistatult mingeid kindlaid koodoneid. Kontekst on see, millised koodonid milliste
naabruses esinevad. Lugemisraami sees ei ole translatsiooni ja transkriptsiooni signaale.
Korduvad järjestused (homopolümeersed järjestused) on alaesindatud, kuna võivad põhjustada
ribosoomi raaminihkeid. Teatud järjestused esinevad väga harva: translatsiooni ja
transkriptsiooni signaalid. Korduvad järjestused on alaesindatud, kuna võivad põhjustada
ribosoomi raaminihkeid
DNA ja RNA koostis
Kõige tähtsam erinevus on suhkrus (suhkru 2’ hapniku olemasolu või puudumine). DNAs
desoksüriboos ja RNAs riboos. DNAs tümidiin ja RNAs uridiin – oluline erinevus, sest uridiini
sattumine DNAsse on DNA reparatsiooni alustamise signaaliks. DNAs on vähe modifikatsiooni
tüüpe, mis piirduvad praktiliselt ainult metüleerimise ja hüdroksümetüleermisega
(metüleeritakse A ja C nukleotiide). RNAs on aga palju erinevaid posttranskriptsioonilisi
modifikatsioone (sünteesijärgseid), mis on keemiliselt väga mitmekesised.
RNA on keemiliselt struktuurilt labiilne ja laguneb väga kergesti 2’ hapniku tõttu. Kui RNA
lähedals satub metalliioon, siis see atakeerib 2’ hapnikuga. DNA on keemiliselt inertne,
stabiilne. 2’ hapniku tõttu on fosfodiesterside RNAs väga labiilne.
DNA on reeglina kaheahelaline, kuid esineb ka üheahelalisi (bakteriofaagid). RNA on ka suurel
määral kaheahelaline, aga see moodustub iseendaga paardumise teel(hairpin struktuur).
Kaksikahela omadused on erinevad neil. DNA kaksikheeliks on reeglina B-vormis, mis on
painduv (saab liigutada rõngasse, topoloogiat on kerge muuta). RNA kaksikheeliks on A-vormis,
mis on väga jäik ja ei paindu. RNA kaksikheeliksil on sageli sees ebaregulaarsused. DNA
struktuuri stabiilsuse tagab eelkõige lämmastikaluste paardumine. RNA struktuuri määrab piki
ahelat aluste sätkumine ning 2’ hapniku ja 4’ hapniku vahelised H-sidemed. DNA on loomult
fibrillaarne ja RNA pigem sarnane valgu struktuurile (globulaarse ehitusega). RNA puhul
aluspaaride vahe piki ahelat väiksem ja seega seonduvad tugevamalt.
DNA
RNA
suhkur 2’desoksüriboos
riboos
(desoksü)tümidiin
(ribo)uridiin
vähe modifikatsiooni tüüp (A ja C metülatsioonid)
sisaldavad sageli erinevaid modifikatsioone (üle
100)
53
DNA koosneb kahest eraldi ahelast
moodustab kaheahelalisi piirkondi paardudes
iseendaga
DNA kaksikheeliks on painduv ja regulaarne
kaksikheeliks jäik, sageli ebaregulaarne
N-aluste paardumisel juhtiv osa
aluste stäkkmine ning 2’O ja 4’O H-sidemed
Struktuur fibrillaarne
globulaarne
tRNA sekundaarstruktuur
Loomult adaptormolekul, mille tasemel realiseerub geneetiline kood. Ühendatakse kovalentselt
AHd kindla nukleotiidse järjestusega, kus AH käib 3’ molekuli otsa (viimase A nukleotiidi otsa).
tRNA pikkus on keskmiselt 76 nukleotiidi. Esimene nukleotiidi on alati G ja viimane 3’ otsas A
(nr 76) – ükskõik kui mitu nukleotiidi tRNAs on). Nt kui tRNAs on 103 nukleotiidid, siis see
viimane A on siiski 76s. tRNAdes milles on üle 80ne nukleotiidid, on variaabel osa väga pikk.
tRNA sekundaarsturktuur ja eriti tertsiaalstruktuur on konserveerunud, mis koosnevad kindla
suurusega struktuursetest elementides (kaksikahela osa on kindla pikkusega).
tRNA struktuur algab aktseptorõlaga, mis aktsepteerib AH. Koosneb alati 7st aluspaarist. RNAs
on võimaliku GU paarid ja UU paarid. D- ja T-õlg, mille nimetused tulenevad lingudest
(üheahelaline ling), mis sisaldab ühte modifitseeritud nukleotiidi – dihüdrouridiin (täpselt nagu
uridiin, aga puudub kaksikside süsinike vahel). Kuna puudub kaksikside, muutuvad omadused
ja mis omakorda muudab lingu omadusi. Dihüdrouridiin moodustab ebastabiilseid aluspaare,
kuna puudub kaksikside. D-õla pikkus võib varieeruda. Antikoodon õlg on alati 5 aluspaari pikk,
mille otsas on antikoodon ling, mis on alati 7 nukleotiidi. Antikoodon lingu otsas asub
antikoodon (3 tk), mis kannavad numbreid 34, 35 ja 36 – paarduvad mRNA koodoniga. T-õlg on
5 aluspaariline, mille otsas on T-ling (tuleneb ribosüüm tümidiinist ehk 5-metüüluriidinist). Kui
tRNA molekul on valmis tehtud, siis lisatakse uridiini 5ndale süsinikule metüül. Pseudouridiin.
Seal, kus üks õlg läheb üle teiseks õlaks, on modifitseeritud lämmastikalused. Just sellised N-
alused, mis ei saa moodustada Watson-Crick aluspaare. Antikoodonis on 1 positsioon (37s) on
hüpermodifitseeritud – see nukleotiid ei saa paarduda (vähemalt W-C paare), lõpetab mRNA ja
tRNA vahelise paardumise. 34 nukleotiidi paardub 3nda nukleotiidiga, kuid on modifitseeritud
nii, et saaks paarduda rohkem kui 1 aluspaariga. tRNA 3’ otsas on universaalselt
konserveerunud CCA järjestus, mis tuntakse ribosoomis ära kohas, kus sünteesitakse
peptiidsidet.
Sekundaarstruktuurist ruumiline struktuur: ruumiline on L-tähe kujuline, milles on praktiliselt
kõik nukleotiidid paardunud (üksikud üheahelalised nukleotiidid vahel, mis ei saa paarduda). D-
ling paardub T-linguga.
tRNA 3’ otsa lisatakse AH, mida kutsutakse aminoatsüleerimiseks, mida viib läbib
aminoatsüülsüntetaasid. Kõigepealt AH aktiveeritakse ensüümi pinnal ATPga, tekib
aminoatsüüladenülaat (saab eksisteerida ainult ensüümi keskkonnas, seal kus vett pole).
Vabaneb pürofosfaat, mis lahkub. Järgmises etapis aminoatsüüladenülaat, mis on kompleksis
ensüümiga, reageerib tRNAga ja AH kantakse üle tRNA 3’ otsa, tekib aminoatsüültRNA.
Vabanevad ensüüm ja AMP.
54
AminoatsüültRNA süntetaase on iga AH jaoks oma ensüüm. 20 AHd seega 20 erinevat ensüümi,
mis jagunevad kahte klassi. Mõlemas klassis on 10 ensüümi. Nad on erineva ehitusega ja
ilmselt erineva evolutsioonilise päritoluga. Klass I ensüümid koosnevad kas 1st subühikust või
2st ühesuguset subühikust. Klass II ensüümid on keerulisema ruumilise struktuuriga.
Klass I ja klass II ensüümid tunnevad ära tRNA molekuli erinevalt. Aminoatsüülsüntetaasid
peavad ära tundma mitmeid tRNAsid. Arginiinil ja leutsiinil on 4 erinevat tRNAd –
aminoatsüülsüntetaas peab tundma need 4 kindlat tRNAd, mis vastavad erinevatele
koodonitele. Klass I ja klass II ensüümid seonduvad tRNAga erinevalt poolt. Need nukleotiidid,
mida aminoatsüültRNAsüntetaasid kasutavad tRNAl ära tundmiseks, nimetatakse tRNA
identsuselementideks. Antikoodon ling on võrdselt kättesaadav mõlemalt poolt.
Klass II – T-ling ja klass I – D-ling.
tRNA identsus
61 koodoni transleerimiseks on enamasti rohkem kui 40 erinevat tRNA liiki. Ühele AHle vastab
1-4 erinevat tRNA molekuli. Ühele AHle vastavad tRNAd sisaldavad samu nukleotiide kindlates
kohtades (nn identsuselemendid). tRNA identsuselemendid on konserveerunud. Nt Ala tRNAdel
63s nukleotiid alati G ja 70s alati U – tekib GU paar, mida ei esine teistel tRNAdel (ainult Ala
tRNAdel). On olemas ka antideterminandid.
Valgusünteesi preribosomaalne etapp – aminoatsüleeriminse reaktsioonid.
2 etappi:
o AH reageerib ATP-ga. AARS – aminoatsüül tRNA süntetaas, see
katalüüsib.
aa+ATP+AARS <-> aaRS~aa-AMP+PPi. Aminoatsüüladenülaat.
o aaRS~aa-AMP + tRNA aatRNA+AARS+AMP. Aminoatsüül tRNA.
Translatsioon
mRNA on kodeeritud tuumas kromosoomides, milles on kodeeritud sünteesitava valgu
järjestus. Valgu sünteesi aparaat on võrdlemisi mahukas ja suur. mRNA järjestust loetakse 5’-3’
suunas ja valku sünteesitakse N-termius→C-terminus suunas. Kõige pealt väljub N-terminaalne
AH, millel alfa-rühm on vaba. Viimane AH on see, mis asub valgu C-terminaalses osas. AHsid
ühendab omavahel ribosoom.
Ribosoom koosneb 2st subühikust (ebavõrdsed), mida nim väikeseks ja suureks subühikuks.
Ribosoomid on oma põhi struktuurilt sarnased kõikides organismides: alati 2 subühikut (väike ja
suur), mis moodustavad tervikliku ribosoomi. Iseloomustatakse Svedbergi sedimentatsiooni
koefitsiendiga, mis on seotud molekulmassiga (mitte lineaarselt). Protsessi, kus ribosoom
seondub mRNA startkoodoniga ja sünteesib valgu valmis ja jõuab üheni 3st stoppkoodnist, nim
ribosoomitsükliks. Ribosoomi subühikud lahutuvad.
Ribosoomil on terve rida funktsionaalseid tsentreid, mis on tRNA sidumiseks (3 kohta), mida
nimetatakse tRNA-sidumissaitideks (A, P ja E sait), kus mRNA seondub väikese subühikuga.
Kasvav peptiid läheb läbi suurema subühiku, AHd ühendatakse suures subühikus. mRNA ja
tRNA vastavusse seadmine toimub väikeses subühikus (P-tsentris). A-sait e aminoatsüül-
tRNAsait – tRNA seondub sinna esmalt. Initsiatsiooni käigus seondub initsiaator tRNA kõigepealt
P-saiti. E-sait on sait, kust sünteesitav valk ribosoomist väljub.
55
Translatsioon jagatakse 3ks etapiks: initsiatsioon, elongatsioon ja terminatsioon. Initsiatsiooni
käigus on kõige olulisem protsess avatud lugemisraami määramine, mis toimub initsiaator-
tRNA abil. Selle käigus seotakse vastavusse initsiaator-tRNA initsiaatorkoodoniga, mis leiab
aset ribosoomi P-saidis.
Initsiatsioon prokarüootides
Initsiatsiooni faktorid: IF1, IF2 ja IF3. IF1 ja IF3 on väikesed valgud (Mw=10 000 Da), u 100
AHd
. IF2 on 10x suurem, koosneb ligikaudu 900 AHst, funktsiooniks on siduda initsiaator-tRNA
ribosoomi väikese subühikuga ning hiljem seonduda initsiaator-tRNA ja initsiaatorkoodoni
suurema subühikuga
. IF3 hoiab ribosoomi subühikud lahus enne kui mRNA ja tRNA seonduvad.
IF1 aitab initsiaator-tRNAd suunata P-saiti, seondub A-saidi piirkonnaga ja blokeerib tRNA
seondumise.
Kui IF3 on seotud väikese subühikuga, siis suurem subühik ei saa seonduda, kuna nende vahel
on molekulaarsed interaktsioonid, mida nim subühikute vahelisteks sildadeks (prokarüootides
12, eukarüootides 15). IF3 takistab sildade tekkimist. Väiksema subühikuga seonduvad IF1 või
IF2, mis on ka G-valk kuna seob GTPd. Ribosoomi etappe katalüüsivad paljud G-valgud. GTPd
siduvad valgud on omavahel sarnase struktuuriga, kus G-domään on sama päritoluga.
Kui ribosoomi väikese subühikuga on seotud mRNA ja IFid, siis selle kompleksiga saab seostuda
initsiaator-tRNA. Võib ka toimuda nii, et IF2 seondub GTPga ja initsiaator-tRNAga väljaspool
väikest subühikut.
Et üles leida õige AUG, kasutab ribosoom lisajärjestust. Kui AUG asub 5’ otsa suunas, kasuta SD
e
Shine Dalgarno e RBS – kui AAGG on AUG järjestusest kindlas kauguses 5’ otsa suunas
(ülesvoolu), siis kokku need 2 signaali moodustavad olulise ja harva ettetuleva järjetuse; sellise,
mis suunab translatsiooni initsiatsiooni õige AUG poole.
Shine Dalgarno funktsioonid: 1) aidata üles leida initsiaator koodon 2) suunata ribosoomi
peatumist e translatsiooni aeglustumist. Järjestuse ära tundmine toimub aluspaardumise alusel:
16S rRNA 3’ otsas viimased kümmekond nukleotiidi on pürimidiini rikas, mis koosneb põhiliselt
U ja C, mis paardub mRNA SD järjestusega.
30S initsiatsiooni kompleks – IF2 aitab 50S subühikul 30Sil seonduda, kuid selleks peab IF3
lahkuma. Kui IF3 lahkub, toimub subühikute omavaheline seondumine ja IF2 seondub ribosoomi
faktorite seondumistsentriga. Piirkond, mis koosneb ribosoomi valkudest ja RNAst. Kui IF2
lahkub, lahkub ka IF1. Selle tulemusena tekib 70S initsiatsiooni kompleks, kus lugemisraam on
paika pandud, kuid on ainult üks tRNA. Vajalik on 2 tRNAd selleks et hoida mRNAd ja
lugemisraami paigal. Lugemisraami säilitamine on valgusünteesi täpsuse tagamiseks väga
oluline.
Initsiatsioon eukarüootides
Eukarüootides on 6 initsiatsiooni faktorit (IF1, IF2, IF3, IF4, IF5 ja IF6). IF4 on eukarüooti
spetsiifiline. IF5B ja IF2 on palju spetsialiseeritum funktsioon kui bakterites oleval IF2-l. IF5A-l on
bakterites olemas analoog elongatsiooni faktorina. eIF1A, eIF3, eIF5B-GTP ja eIF2-Met-tRNA
aitavad initsiaator-tRNAl seostuda ribosoomiga. eIF4E seondub mRNA 5’ otsaga ehk mis on RNA
polümeraas II produktid, mis võib toimuda kas tuumas või tsütoplasmas. eIF4G ja temaga
seotud valkudega seonduvad mRNAga ribosoomist sõltumatult.
40S pre-initsiatsiooni
kompleks ja eIF4 seonduvad omavahel. 40S pre-initsiatsiooni kompleks koos Met-IF4
56
subühikutega kokku moodustavad 48S initsiatsiooni kompleksi, hakkab liikuma mööda mRNAd
– mRNA skaneerimine, mida skaneeritakse initsiaatorkoodoni suhtes. Kui initsiaator-koodon on
üles leitud, siis saab seonduda ribosoomi suurem subühik ja eIF2 ja eIF5l toimub GTP hüdrolüüs,
moodustub 80S initsiatsiooni kompleks.
AUG koodon peab olema õiges kontekstis, et saaks alata translatsioon. Õige kontekst tähendab
seda, et raamistavad nukleotiidid peavad olema sobivad. Kõige olulisem positsioon on -4 (4
koodonit enne AUGd), kus peab olema A koodon. Kozaki konsensus
.
RNA helikaas – suunab RNA kaheahelalise struktuuri lahti harutamist. Selleks et mRNa
skaneerimine saaks edukalt toimuda on vaja IF4A kompleksis IF4Bga – et ribosoom saaks
liikuda mööda mRNAd.
Elongatsioon
Osalevad ribsoomiga seotud elongatsiooni faktorid, mille käigus toimub aminoatsüül-tRNA välja
valimine ribosoomi A-saidis, milles osaleb EF-Tu. Elongatsiooni käigus toimub peptiidsidemete
süntees. Toimub translokatsioon, mida aitab läbi viia EF-Tu ja EF-G. Aminoatsüül-tRNA on
kompleksi EF-Tuga, mis on seotud aktseptor õlaga. See kompleks seondub ribosoomi A-saiti nii,
et EF-Tu on AHga seotud, mis ei saa osaleda peptiidsideme moodustumisel nii kaua kui EF-Tu
on seal. Kui leitakse aminoatsüül-tRNA, mille antikoodon paardub mRNA koodoniga
(stohhastiline otsimine), siis ribosoom moodustab H-sidemeid koodon-antikoodon kompleksiga.
Moodustub stabiilne kompleks.
Väikese vao poolt kontrollitakse aluspaardumise geomeetriat – kui ei ole W-C paardumine, siis
lisa H-sidemeid ei moodustu ja tRNA ei seondu stabiilselt ribosoomi A-saiti. Kui toimub
paardumine, siis EF-Tu interakteerub faktorite sidumiskeskusega, mis aktiveerib EF-Tu GTP
hüdrolüüsi, EF-Tu lahkub ja aminoatsüül 3’ ots saab ribosoomis akumuleeruda ja seonduda
ribosoomi sellesse tsentrisse, kus toimub peptiidsideme moodustumine. Kasvav peptiid
kantakse üle A-saidis oleva aminoatsüül-tRNA külge, tekib uus peptidüül-tRNA, mis on ühe AH
võrra pikem (kasvab N-C suunas). Peptiidi ülekande tõttu ribosoomi peptiid sünteesivat kohta
nim peptidüül- transferaasne tsenter. Katalüüsib reeglina ainult ribosoom ise, kuna ei vaja lisa
faktoreid peptiidsideme sünteesiks, v.a. proliin (kuna on iminohape). Polüproliin järjetuse
sünteesiks on vaja lisafaktoreid, mida nimetatakse bakterites EF-P ja eukarüootide eIF5A.
Tekib olukord kus A-saidis on peptidüül-tRNA ja P-saidis transleeritud tRNA ning et saaks uus
tRNA lisanduda on vaja lükata eemale teised – translokatsioon. EF-G katalüüsib 2 tRNA ja mRNA
liikumist ühe koodoni võrra edasi. 2 järjestikust koodonit on kompleksis mRNAga – 6 aluspaari
abil hoitakse mRNAd paigal ja seega lugemisraam on paigal. EF-G ja EF-Tu kompleksis GDPga
on inaktiivne. Nukleotiidid vahetuseks on vaja lisafaktorit. EF-Tu kompleksis GTPga + EF-Ts??
EF-G ei vaja lisafaktorit, kuna saab nukleotiidi vahetust toimetada ise, seob GTPd ja GDPd
enam-vähem võrdselt.
Terminatsioon
Universaalses koodis on 3 UAG, UGA ja UAA. Terminatsiooni koodoneid tunnevad ära
terminatsiooni faktorid, mida on kahte klassi. Klass I kuuluvad bakterites RF1 ja RF2,
57
eukarüootides eRF1. Klass II kuuluvad bakterites RF3 ja eukarüootides eRF3
. Klass I faktorid täidavad ribosoomis tRNA funktsiooni ehk tunnevad ära mRNA. Peptiidne-antikoodon. RF1
tunneb ära UAG, RF2 UGA ja RF1+RF2 UAA. UAA on väga tugev stopp-koodon. UAG ja eriti UGA
koodon on kõige sagedasem stopp-koodoni üleminek. Stopp-koodoni ülelugemine on oluline.
RF1 ja RF2 seondub A-saiti, kui seal on eksponeeritud üks kolmest stopp-koodonist. RF1 ja RF2
on struktuurilt sarnased tRNA valgud, samasuguse kujuga ja seonduvad A-saiti. Nende ühte
osa, mida nim peptiidseks antikoodoniks tunneb ära mRNA. GGQ katalüüsib peptiidahela
vabanemist (glütsiin-glütsiin-glutamiin järjestus). Ribosoom on endiselt kompleksis RF1 ja
RF2ga. RF3 on kompleksis GTPga, mis on ebatavaline nähtus, kuna ta seondub ribosoomiga
GDP vormis. Kui kasvav polüpeptiid on lahkunud, toimub RF3el GTP vahendus, kus GDP
vahetatakse GTP vastu – G-nukleotiidi vahetusfaktor. Selle protsessi käigus klass I
terminatsiooni faktor lahkub. GTP hüdrolüüs suunab RF3 lahkumist. 2 tRNAd, mRNA ja
ribosoomi mõlemad subühikud on ühes kompleksis – selline ribosoom ei saa alustada uut valgu
sünteesi enne kui tRNA ja mRNA on välja vahetatud e on vabastatud neist. 2 tRNAd hoiavad
mRNad kinni, selles osalevad lisaks RRFile (ribosoomi vabastamise faktor), IF3 ja EF-G -
suunavad ribsoomi retsükleerimist. RRF seondub ribosoomi A-saiti, seejärel seostub EF-G, mis
katalüüsib translokatsiooni (mRNA ja RRF liikumist ribosoomis, lükkab ribosoomist välja
deatsüleeritud tRNAd). Väikse subühikuga seostub IF3, mis muudab protsessi pöördumatuks.
Ribosoomi struktuur
Avastati 50ndatel, leiti ERi pinnalt. Ribsoomid on organiseeritud polüsoomidesse. Translatsiooni
lõpus ribosoomi subühikud dissotseeruvad. Liigub 5’3’
Ribosoomid koosnevad RNAst ja valkudest. Katalüütilised tsetrid koosnevad RNAst. Ribosoomid
on ribosüümid. rRNA on katalüütiline RNA. Ribosoomi aktiivtsentris on ainult RNA, sinna
lähedusse ulatuvad ka mõned valgud. Peptiidsideme sünteesi eest vastutav tsenter koosneb ka
RNAst. Valgud, mis sinna ulatuvad on stabiliseeriva, mitte katalüütilise funktsiooniga.
Valgu sünteesi katalüüsib RNA. Elu tekke alguses oli RNA, mis hakkas katalüüsima iseenda
replikatsiooni. Siis hakkas RNA sünteesima ka valke. Alates programmeeritud valgu sünteesi
tekkest räägime elust.
Peptidüül-tRNA katalüüsib peptiidsideme sünteesi. Enamus biokeemilisi reaktsioone
katalüüsivad valgulised ensüümid, valkude enda sünteesi aga RNA.
Ribosoomid on väga konserveerunud oma struktuurilt. Võivad küll olla erineva suurusega, kuid
kõik tsentrid on väga sarnase ehitusega.
Ribosoomi komponendid
Prokarüoot
Eukarüoot
Suurus
70S 2.5MDa
80S 4MDa
Subühikud
30S+50S
40S+60S
rRNA
16S (1500 nt)
23S (2900 nt)
18S (2000 nt)
28S (4000 nt)
58
5S (120 nt)
(70%)
5,8S (160 nt)
5s (120 nt)
(60%)
r-valgud
55 (30%)
21+34
80 (40%)
33+47
5S RNA on homoloogne prokarüoodis ja eukarüoodis. Eukarüootse ribosoomi mass on suurem
valkude arvelt valdavalt.
Svedbergi koefitsient: Ribosoome lahutatakse tsentrifuugimisega: rakendatakse proovile
raskusvälja, 100 000 või rohkem korda suurem, kui maa raskusjõud. Raskusjõu mõjul erineva
suuruse ja tihedusega partiklid liiguvad erinevatesse kihtidesse. Kõige tihedamad on
rakutuumad. Kõige kiiremini liiguvad terved ribosoomid 70S, siis suur subühik 50S ja siis väike
30S.
Ribosoomi struktuurne organisatsioon:
•
Ribosoom on globulaarse struktuuriga mille keskne komponent on ribosoomi RNA
(rRNA), mis täidab ribosoomi põhilisi funktsioone.
•
rRNA koosneb põhiliselt paardunud osadest (heeliksitest), mis on seotud
paardumata osadega.
•
rRNA moodustab struktuurseid domeene, mis on r-valkudega seotud jäikadeks ja
monoliitseks üksusteks
•
Ribosoomi valgud (r-valgud) seonduvad rRNAga ja stabiliseerivad selle struktuuri
•
r-valgud on funktsionaalselt olulised ja täidavad mõningaid spetsiifilisi ülesandeid
(mRNA sidumine, peptiidi väljumine, faktorite sidumine)
•
Paljud r-valgud sisaldavad globulaarset domeeni ja pikka “üheahelalist saba”, mis
seondub rRNA’ga.
•
rRNA on enam konserveerunud struktuuriga
•
r-valgud on osaliselt kattuvad eri riikide ribosoomidel
•
Mõlemad on funktsionaalselt olulised
•
Ribosoomid on globulaarsed. rRNA moodustab ribosoomi karkassi, keskse struktuuri. rRNA
vahelised augud täidavad ribosoomi valgud. rRNA koosneb väga paljudest suhteliselt
lühikestest kaksikahelalistest osadest, heeliksitest. RNA-RNA interaktsioonid on põhilised, mis
hoiavad ribosoomi karkassi koos.
rRNA molekulid moodustavad iseseisvaid struktuurseid domääne. Neid domääne stabiliseerivad
ja nende vahelisi interaktsioone vahendavad ribosoomi valgud. Ribosoomi valkudel on väga
oluline osa ka katalüütilistel protsessidel, annavad ribosoomile dünaamika, ribosoomi struktuur
saab muutuda ühest vormist teise kiirelt tänu ribosoomi valkudele.
Ribosoomi valgud erinevad teistest valkudest, sest neil on erakordselt pikad üheahelalised
sabad, ligi pool AH-lisest järjestusest. Need sabad lähevad RNA heeliksite vahele ning
ühendavad erinevaid RNA osi.
59
rRNA on evolutsiooniliselt väga konserveerunud. Tema järjestuse alusel saab moodustada
evolutsiooni puu.
Ribosoomid on omavahel seotud paljude subühikute vaheliste sildadega. On RNA-RNA sillad ja
valgulise kompnendiga sillad(need pigem perifeersed). Bakteril on 12 silda.
Ribosoomi subühikud liiguvad üksteise suhtes translatsiooni käigus.
Väike subühik koosneb 3 erinevast domäänist, mis võivad üksteise suhtes liikuda. Suurem
subühik on jäiga struktuuriga, 6 domääni on omavahel seotud. Dünaamika annavad ribosoomile
ribosoomi valgud, valgud võivad liikuda ühelt ribosoomilt teisele või vahetuda lahuse ja
ribosoomi vahel.
Ribosoomi subühikud on veidi erineva organisatsiooniga:
•
Väike subühik koosneb kolmest eraldi domeenist, mis võivad üksteise suhtes
liikuda.
•
Suurem subühik on monoliitselt jäiga stuktuuriga.
•
R-valgud on dünaamilise struktuuriga ja mõned neist võivad “ära käia”.
•
Enamus r-valke on ribosoomis vahetatavad
•
Aktiivtsentrid – väiksema subühiku pea ja platvormi ahel asub dekodeeriv
tsenter.
•
mRNA kanal kulgeb mööda väiksema subühiku kaela
•
Kasvav peptiidi tunnel
Väike subühik: 3’ domään, 5’ domään ja keskmine domään. Heeliks 44 kontakteerub
kõigi domäänidega.
Suur subühik: monoliitsema organisatsiooniga, 6 struktuurset domääni: 5S RNA
domäänis asuvad paigad, mis osalevad peptiidsideme sünteesil.
rRNAs on ekstensioonisegmendid – RNA lisaheeliksid ja lingud molekuli keskel.
Lisasegmendid jäävad perifeersesse alasse välja poole.
Mutatsioonid valkudes muudavad translatsioonitäpsust
. S4 ja S5 reguleerivad seda, kui
tugev on koodon-antikoodon interaktsioon ehk kui tugevalt nad koos püsivad
(kontrollivad ribosoomi töö täpsust). Mutatsioonid nendes valkudes põhjustavad
translatsiooni vigade sagedust.
Translatsiooni kiirus
: Translatsiooni initsiatsioon ja terminatsioon on suht aeglased, võtab aega
1-5 sek. Bakteri ribosoomid suudavad valku sünteesida väga kiiresti
10-20 AH sekundis.
Translatsioon on kodeeritud transkriptsiooniga, valgu süntees toimub sama kiiresti kui mRNA
süntees. Eukarüootide elongatsiooni kiirus on väiksem kui bakteritel,
2-4 AH sekundis.
Translatsiooni täpsusest:
o Keskmine vigade sagedus bakterite valgusünteesil on vähem kui
1/1000 vale aminohappe lülitumise jaoks.
o Stop koodonite ära tundmine on erineva täpsusega – UAA ära
tundmine on täpseim(RF1, RF2). UGA tunneb ära vaid RF2, selle
60
äratundmine on suhteliselt vähem täpne ja UGA kohal esineb sellina
nähtus, mida kutsutakse read-through translatsiooniks.
o Raaminihke vigade sagedus on väiksem, kui 10^(-5)
Antibiootikumid ja valgusüntees
Ribosoomiga seonduvad antibakteriaalsed ained. Inimese poolt kasutatud antibiootikumid on
keemiliselt sünteesitud. Enamike antibiootikumide märklaud on valkude süntees. Paljud
seonduvad ribosoomiga ja toimivad ribosoomi tasemel.
Antibiootikumide (penitsiliini) avastaja
Alexander Fleming 1929.
Vankomütsiin tekitab bakteri kesta sisse auke.
Sulfonüülamiidid pidurdavad
bakterirakkudes foliinhappe biosünteesi (sisaldub värvaines).
Aminoglükosiidid
Suhkruderivaadid, mis koosnevad suhkrujääkidest, mis on omavahel seotud
glükosiidsidemetega. Suhkrud sisaldavad aminorühmasid kindlates kohtades
. Kanamütsiin seondub ribosoomi dekodeerivasse tsentrisse. Suurendab valgusünteesi vigade sagedust kuni
100x. Kui A-saiti satub vale tRNA, ei hoita mRNAd nii täpselt kinni, lugemisraam nihkub,
sünteesitakse ainult väga lühikesi valke, mis indutseerib proteaase. Proteaaside väga kõrge
tase tingib ka õigete valkude lagundamise ja rakk sureb lõpuks. Aminoglükosiidid takistavad ka
translokatsiooni.
•
Suurendavad valgusünteesi vigade sagedust (norm 1/1000-10000)
•
Str, neo puhul vigade sagedus kuni 100x kõrgem, mis viib translatsiooni
peatumiseni raaminihke tõttu
•
Inhibeerivad aa-tRNA seondusmist ribosoomi A-saiti (Str, kana)
•
Takistavad ribosoomi translokatsiooni (Spc, Neo)
Tetratsükliin
Normaalse translatsiooni korral seondub EF-Tu:GTP kompleks, antikoodoni järgi valitakse välja
tRNA, toimub stabiilne seondumine, GTP hüdrolüüs, EF-Tu lahkub, A sait jääb seotud
aminoatsüül-tRNAga.
Kui tetratsükliin seostub, siis algset tRNA
valikut ta ei sega, aga stabiilset seondumist ei
teki. EF-Tu ei teki GTP hüdrolüüsi ning see
kolmikkompleks lahkub ribosoomilt.
Tetratsükliin takistab aminoatsüül-tRNA
seondumist ribosoomi saiti.
Tetratsükliinil on ribosoomi sidumiskohti väga
palju.
Tigetsükliin keemiliselt veits modifitseeritud.
Seondub sarnaselt, kuid mõned rühmad on
61
natuke teises asendis või rohkem. Tetratsükliini resistentsed bakterid on tundlikud
tigetsükliinile.
Puromütsiin – sarnane aminoatsüül-tRNA 3’ otsaga. Lämmastik muudab keemiliselt
stabiilseks. Kui peptiid kantakse üle puromütsiinile, siis ta ei jää ribosoomi pidama, vaid
dissotseerub ribosoomilt, valgu ahelat ei sünteesita edasi enam. | Ta on väga sarnane osaliselt
aminoatsüül-tRNA-le, ta seondub sinna, kus toimub peptiidsidmete tekkimine. Valk
sünteesitakse kuni puromütsiinini ja edasi ei lähe. Poolik valk on rakule ju ometi halp.
Kloramfenikool
Väike aromaatne molekul
. Bakterispetsiifiline AB, ei takista valgu sünteesi eukarüootidel.
Seondub ribosoomi peptiidsidet katalüüsivasse tsentrisse. Spetsiifiliselt inhibeerib aromaatse
külgahelaga AHde liitumist valkudesse. Takistab peptiidsideme sünteesi ja aromaatsete
aminohapetega aa-tRNA sidumist ribosoomi A-saiti.
Makroliidid
Takistavad kasvava peptiidi liikumist ribosoomi sees. Seonduvad tunneli kitsaste kohtade
peale. Takistavad kasvava peptiidi sisenemist tunnelisse, translatsioon seiskub kui peptiidi ei
saa pikendada.
Antibiootikumide resistentsuse tagavad erinevad teed
AB resistentsed bakteri tüved tekivad enamasti haiglates, kus ABsid palju kasutatakse.
Multiresistentsed tuberkuloosi tüved on päris vanglatest. Mycobacterium tuberculosis.
AB märklaua modifitseerimine – ribosoom on AB märklaud, kui seal modifitseerida ära
need keemilised rühmad, mis peaksid ABga H-sidemeid tekitama, siis AB enam ei soendu
ja ribosoomid muutuvad AB resistentseks.
AB modifitseerimine – atsetüleerides nt kloamfenikooli jms Absid, et märklauaga
seondumist ei toimuks
AB rakust välja pumpamine – peab toimima kahe eelmisega koos.
Mutatsioonid ribosoomides tagavad resistentsuse ABde suhtes
Streptomütsiin – (seondub ribosoomi dekodeerivasse tsentrisse) mutatsioonid valgus S12 ja
16S rRNA nukleotiidides. Ei seondu mutantsete ribosoomidega enam.
Makroliidid – nt erütromütsiin, mutatsioonid vallkudes L4 ja L22 ja 23S rRNAs. 23S rRNAs
takistavad erütromütsiini seondumist otseselt ribosoomidega. L4 ja L22 ei mõjuta makroliidide
seondumist ribosoomidega, aga muudavad ribosoomi tunneli geomeetriat.
Kloramfenikool – mutatsioonid 23S rRNAs, lisaks cat geeenis.
Eukarüootide spetsiifilised ABd on ka olemas, bakteri ribosoomile nad ei mõju. Bakteri ABd ei
satu eukarüoodi rakku, eukarüoodi rakumembraan ei ole läbitav bakteri ABle, st bakteri ABd ei
mõjuta eukarüoote tavaliselt. Bakterid tõmbavad ise sisse ABsid. Nt tetratsükliin inhibeerib, aga
ei lähe rakku sisse.
Tetratsükliin inhibeerib eukarüoote, aga ei lähe rakku sisse.
Tsükloheksimiid blokeerib translokatsiooni eukarüootidel, kuid baktereid ei mõjuta.
62
tmRNA
transport-messenger RNA, nii tRNA kui mRNA omadused, aminoatsüleeritakse alaniiniga, aga
sisaldab ka kodeerivat osa (11 koodonit). Sisaldab mõlemat tRNA ja matriitsRNA osi. Sarnane
alaniini tRNAga üks osa temast. Seondub sellise ribosoomiga, kus translatsioon on mingil
põhjusel seisma jäänud. Tekib surnud kompleks, tuleb mängu tmRNA, päästab ribosoomi
blokist. tmRNA osaleb katkenud mRNA translatsiooniproduktide lagundamisse suunamisel
Ribosoom, mis mingil põhjusel edasi ei sünteesi, nt AB inhibeerib või mRNa on katkenud.
tmRNA, mille 3’ otsas on alaniin, seostub ribosoomi suurem subühiku A saiti. tmRNA on on
kompleksis ühe EF-Tu sarnase valguga, mis aitab tal ribosoomiga seonduda. Kodeerib 11 kindla
koodoniga AHd. Nüüd aga on valgu C-terminaalses osas lisaks see alaniin(kodeerimata) ja 11
AH-d, mis on signaaliks proteaasidele valk lagundamisele.
Bakterite ja eukarüootide translatsiooni võrdlus
Bakter
Eukarüoot
RNA ja valgusüntees seotud, teineteist toetavad
(transkriptsioon jääb seisma, kui sellelt mRNAlt ei
toimu valgu sünteesi)
RNA süntees lahutatud valgusünteesist,
sõltumatud
(transkriptsioon tuumas ja translatsioon
tsütoplasmas)
mRNA polütsistroonne
(üks mRNA sisaldab infot mitme valgu sünteesiks)
mRNA monotsistroonne
initsiatsioon tänu SD järjestusele
AUG kontekst
Translatsioon kiirem 10-20 AH/sek
Translatsioon aeglasem 2-4 AH/sek
Bakterites translatsioon:
o Transkriptsioon ja translatsioon on üheaegsed ja üksteist toetavad.
Kui ribosoom transleerib valku, siis RNA pol transkribeerib
kindlamalt.
o mRNA on polütsistroonne – ühe mRNA pealt mitu eri valku.
o Initsiaatorpiirkonnas on lisaks AUG koodonile ka SD järjestus, 4-8nt
järjestus, mis paardub 16sRNA-ga, ta on AUG ülesvoolu – 5’ otsa
suunas.
o Kiirem protsess.
Eukarüootides:
o RNA süntees lahus valgusünteesist. RNA pol II produktidel 5’ otsas
cap struktuur, neid splaissitakse ning kompleksis mitmesuguste
valkude viiakse tsütoplasmasse – toimub palju sündmusi enne, kui
saab tsütoplasmas translatsioon toimuda.
o mRNA on monotsistroonsed, kuigi esineb uORF-e.
o Oluline on AUG kontekst – naabernukleotiidid mis AUG-d
raamistavad.
o Aeglasem protsess.
Standardne eukarüootne mRNA.
o 5’ otsas sisaldab cap struktuuri.
63
o AUG koodonit ümbritsevad kindlad nukleotiidid – olulisim järjestus
on gccRccAUGG.(puriin – R). Kozak’i konsensuzjärjestus – avastaja
järgi nim.
o 3’ otsas on ka mitmetransleeriv ala – polyA saba.
Translatsiooni initsiatsiooni ettevalmistamine:
o Eelmise translatsiooni järel jääb alles 40S kompleks, kus küljes eIF3.
o Selle külge eIF5B ja eIF2 ja initsiaator tRNA.
o Erinevalt bakteritest toimub ka mRNA ettevalmistus. Temaga
seonduvad eIF4 initsaator faktorid. Olulisim on eIF4G, on suur
adaptorvalk, millega seonduvad teised initsiatsioonifaktor 4-d.
eIF4B, eIF4E on cap binding, koos nad helikaasi kompleks. Lisaks
tarvis ka polyA binding protein. Translatsiooni initsiatsioonil on
mRNA 5’ cap ja 3’ polyA saba koos.
o Initsiaator tRNA ettevalmistamine. See on metioniini tRNA, millel on
teatud kindel järjestuse struktuurne omapära, mis ei lase tal
elongaator tRNA-na toimida. Ta seondub eIF2-ga, eIF2 koos GTP-ga
ta seondab ja selline kompleks liigub edasi ribosoomi väiksemasse
subühikusse.
o 43S kompleksile(see kus kogu eelnev kompott koos) seondumine
mRNA-le.
o 40S subühik liigub mööda mRNA-d ja initsiaator tRNA abil otsitakse
AUG koodonit – skaneeritakse mRNA-d AUG koodoni suhtes. Kui AUG
üles leitakse, siis tekib tRNA paardumine mRNA-ga ja siis on
lgemisraam edukalt üles leitud. Nüüd toimub eIF2-l GTP hüdrolüüs
ning kompleks, mis jääb(koos mRNA-ga) on 48S kompleks.
o eIF5B-l toimub GTP hüdrolüüs, eIF-d vabanevad ja 60S subühik saab
ka ühineda. Ja translatsioon võib alata edukalt leitud lugemisraami
pealt!
RNA kvaliteedi kontroll päristuumsetes
•
Kui RNA ei täida rakus oma funktsiooni, siis ta lagundatakse
•
mRNA, mis sisaldab stop koodonit enne viimast eksonit või millel puudub stop
koodon või millel jääb translatsioon seisma, lagundatakse
•
rRNA, mis ei moodusta aktiivseid ribosoome (modifitseerimise või protsessimise
defektid), lagundatakse
Nonsense mediated decay
Toimib mRNAl. Kui mRNAl on stoppkoodon enne viimast eksonit ehk siis upf valgud
jäävad viimase ekson-ekson ühendusega seotuks, ribosoom transleerib ja ribosoom ei
lükka upf-e maha. Need upf-id mis jäävad seotuks, kutsuvad kohale eksoribonukleaaside
kompleksi, mis lagundab mRNA ära, millega upf valgud seotud on. Upf valgus
singaliseerivad eksonukleaasidele, et see mRNA ei ole kvaliteetne. Sarnane mehhanism
toimib ka siis, kui mRNAl ei ole stopp-koodonit. Seal ei osale upf valgud, aga on valgus,
64
mis tunnevad ära et sel mRNAl puudub stopp-koodon ja mRNA lagundatakse 3’-5’
suunas.
Upf valgus seostuvad selle piirkonnaga, mis jäävad ekson-ekson ühendusest 50 aluspaari
ülespoole. Kui stopp-koodon on peale viimast ekson-ekson ühendust, siis NMD ei puutu
sellist mRNAd.
Translatsiooniline kontroll
Pre-mRNA läbib splaissimise ja selle splaiss-saidiga (ekson-ekson ühenduse piirkonnaga)
jääb seotuks kompleks, kus on upf valgud jt. Sellises kompleksis transporditakse mRNA
tuumast tsütoplasmasse, kus hakkab seda transleerima ribosoom. Kui ribosoom
transleerib ja midagi ei ole viga, siis lükkab ribosoom upf-e maha, mis lähevad tagasi
tuuma koos eksportiiniga. Eksportiin seostub mRNAga.
Ebakvaliteetse mRNA lagundamine võib toimuda mõlemast otsast. Selleks, et seda 5’
otsast lagundada, tuleb eemaldada cap-stuktuur. 3’ otsa jaoks on vaja polü-A saba
eemaldada.
Esimest translatsiooni tsüklit kutsutakse biomeerseks translatsiooni tsükliks. Selle käigus
eemaldatakse mRNAga seondunud valgud, mis olid seondunud juba tuumas.
No-go decay
Kui mRNA on läinud sõlme, siis selle seisma jäänud sõlmilise ribosoom-mRNA kompleksi
tunnevad ära Hbs1p nt. mRNA lõigatakse keskelt katki ja hakatakse lagundama 3’-5’
suunas ja ka 5’-3’ suunas. 3’-5’ suunas lagundamine on põhiline viis kuidas enamik
RNAsid lagundatakse, mida viib läbi eksosoom (eksonukleaaside kompleks).
NMD vahendatud upf-ide poolt (mRNA lagundamine keskelt mõlemas suunas), NSD Ski7p
poolt (mRNA lagundamine 3’ otsast) ja NGD Hbs1p poolt (lagundamine 5’ ja 3’ otsast).
Transkriptsiooni aktivaator GCN4 norm rakus
o Ta on RNA sünteesi reguleeriv faktor, mis kontrollib metaboolsete
operonide transkriptsiooni. AH sünteesi geenidelt ja mitmesuguste
metaboolsete ensüümide sünteesi reguleeriv.
o Kui rakkudel nt süsinikunälg, siis GCN4 aktiveeritakse. Ning tema
transkribeerimine on ka vaid siis aktiivne. Toimub vaid siis, sest
GCN4 järjestuse ees on 4 uORF(upstream open reading-frames),
need on sellised lühikesed lugemisraamid, mida pole muuks tarvis,
vaid ongi et kontrollida, kas on vaja sünteesida seda nende järgset
produkti või mitte.
Kui on kõik korras rakus, siis sünteesitakse nende pealt lihtsalt
mingid peptiidid ning sellega asi lõppeb – GCN4 ei transleerita.
o Kui rakk nälg, siis uORF-id muutuvad tagasihoidlikumas. Sest eIF2
fosforüülitakse ning on madala energiatasemga. Ja kui eIF2 tase on
madal, siis nende nelja uORF pealt initsiatsiooni ei toimu. Ning see
omakorda suunab ribosoomid GCN4, õige valgu, initsiatsiooni
koodonile.
65
Ko-translatsiooniline transport
Universaalne nagu valgu sünteeski. Toimub kõikidel organismirühmadel (bakterid, arhed,
eukarüoodid). Samal ajal kui ribosoom valku sünteesib, mis on kinnitunud membraanile, valk
läheb kohe otsemat teed läbi membraan (sünteesitakse läbi membraan). Eukarüootide puhul
tsüotplasma luumenisse, bakterite puhul periplasmasse või teistesse membraansetesse
moodustistesse. Universaalne komponent on valk Sec61 (61 kDa, sec - sekretsioonisüsteem),
mis moodustab poori, mille kaudu valk läheb läbi membraani. Teised transpordi liigid esinevad
nii bakteritel ja arhedel – posttranslatsiooniline transport – kui valk on valmis sünteesitud.
Valk läheb läbi ribosoomi suures subühikus oleva tunneli ja see järel läbib
tsütoplasmavõrgustiku membraani, kus asub poor (Sec61). Läbi Sec61 valk läheb
tsütoplasmavõrgustiku valendiku poole. Kui ribosoomi pole, siis pooril on olemas „kork“, mida
nimetatakse valgu translokatsioon
iks (mitte segi ajada ribosoomi translokatsiooniga), mis
tähendab et valk läheb ühelt membraani poolelt teisele. See, kuhu valk sünteesitakse (kas
niimoodi, et jääb tsütoplasmasse ringi käima või läheb läbi membraani), määrab valgu N-
terminaalne järjestus.
Bloebel – pani esimesena tähele, et nendel valkudel, mis suunatakse
tsütoplasmavõrgustikku, on N-terminaalses otsas sarnane järjestus. Esimeses 10 AHs on
vähemalt 6 AHd hüdrofoobse kõrvalahelaga, mis on valgu translokatsiooni signaaliks. See
signaliseerib valgu transportimist ERi.
Rakkudes on 2 kompleksi, mis selle signaale ära tunnevad. Need valgud, millel on N-terminuses
hüdrofoobne signaal, nende ribosoomidega seondub SRP kompleks.
SRP e signaali
äratundev partikkel, mis koosneb 7S RNAst ja 5st valgust. Juhul kui valgu N-terminaalne
järjestus ei ole hüdrofoobne vaid hüdrofiilne, siis selliste ribosoomidega seondub
NAC. NAC
koosneb ainult valkudest. Takistab SRP seondumist.
Ribosoomi kasvava polüpeptiidi tunnelis on 40 AHd. Kasvava polüpeptiid „pistab välja“ pea
tunnelist, siis kui on sünteesitud 40 AHd.
SRP seondub sellise peptiidiga, mis kasvab välja tunnelist. Kui SRP seondub, siis ta indutseerib
translatsiooni peatumist. Kasvab kõiki kvaliteedikontrolli seondujaid ribosoomile seondumast,
et nad ei lagundaks ribosoomi jne laiali. Polüpeptiid liigub nii kaua tsütoplasmas ringi, kuni
jõuab SRP retseptorini, mis asub kareda ERi pinnal. Kui ribsoom on SRP retseptoriga seotud,
jääb ta seotuks ERiga. Toimub lateraalne difusioon mööda tsütoplasmavõrgustiku membraani,
kuni jõuab Sec61 poorini, mis on korgiga kinni (omaette valk). Ribosoom seondub Sec61
pooriga ja SRP ja SRP retseptor aitavad juhatada kasvava polüpetiidid N-terminaalse otsa Sec61
„auku“. Kui valk on poori sisse läinud: kork tuleb ära, SRP koos SRP retseptoriga lahkuvad
tsütoplasmasse. On tekitatud situatsioon, kus ribosoom saab translatsiooni jätkata, aga
ribosoom on seotud kareda ERiga. N-terminaalne signaalpeptiid jääb seotuks Sec61ga ja Sec61
küljest lõigatakse ta lahti. Valmis saanud valk ei jää võrgustikus üksi, vaid selle valguga
seonduvad mitmesugused komponendid, mille hulgas on chaperone.
Valgu voltumine
Kui valk on valmis, siis see pikk polüpeptiidahel hakkab võnkuma. Kuna ta on veekeskkonnas,
siis surutakse valk kokku (eriti hüdrofoobsed osad, mis hakkavad agregeeruma). Selgroos
tekivad H-sidemed. Translatsiooni käigus tekib valgu ruumiline struktuur. Valgu modifikatsiooni
tehakse Golgi kompleksis.
66
Kui valgud on ERis, siis ei tohi nad olla aktiivsed. Need valgud, mis sünteesitakse ERi,
seonduvad chaperonidega, mis ei lase valgu struktuuril muutuda enne kui nad on jõudnud
lokalisatsiooni lõppu. ERis toimub valkude aktiivne pakkimine, struktureerivad, pakitakse ka
omavahel kokku.
Valgu struktuuri moodustumine toimub translatsiooni ajal mitmes järgus. Kui N-terminaalne
domään on ribosoomist välja ulatunud, siis selle koha peal ribosoom jääb sageli seisma, mida
suunavad signaalid (transleeritakse sellised koodonid, mida sünteesitakse aeglaselt).
Translatsiooni seismajäämine ei toimu domäänide piiril, vaid on 40 lisa AHd, mis ei ole
struktureerinud (põhilisedl alfa-heeliks). Kui kogu valk valmis saab, siis saab moodustuda ka C-
terminaalne domään. Selle koha peal, kus kasvav polüpeptiid ribosoomist välja tuleb, on
platvorm, millega seonduvad ribosoomi seoselised chaperonid. Alfa-heeliks läheb läbi suurema
subühiku.
Ribosoom sünteesib, valk pressitakse välja. Valguga seostuvad chaperonid, mis takistavad
valgu struktuuri moodustumist. Valgud võivad minna läbi membraani (posttranslatsiooniline).
Peale sünteesi, valk koos chaperonidega struktureerub. Valk võib agregeeruda teiste
valkudega, ära laguneda. Suur osa sünteesitavatest valkudest lagundatakse enne kui neist
üldse asja saab.
Kui valgu struktuur pole õige või on agregeerunud, läheb degradatsiooni ja muudetakse tagasi
AHdeks.
Chaperon
Hsp 60 suunatud voltumine
Valkude ruumiline struktuur on valkude funktsioneerimise alus. Ruumiline struktuur võib
muutuda siis kui nt temperatuur tõuseb.
Kuuma ehmatuse valgud on need, mida rakk hakkab siis sünteesima kui temp ootamatult
tõuseb. Sellega kaitsevad nad valke temp tõusu eest, taastavad nende ruumilise struktuuri
(temp tõusu tõttu lähevad hüdrofoobsed osad välja poole)
Hsp jaotatakse perekondadesse suuruse järgi. Neid on hästi palju erinevaid. Valgud, mida
sünteesitakse siis kui temp on liiga kõrge, mis aitavad taastada olemasolevate valkude
natiivset struktuuri on Hsp 60, mis käib koos Hsp 15ga. Hsp 60 on chaperon, mis moodustab
rakkude iseloomulikke struktuure. Koosneb 14st monomeerist, mis on organiseeritud 2ks
ringiks, mis on omavahel koos. Kui üks ots on lahti, siis võtab ta sisse valku (mille hüdrofoobsed
osad on suunatud välja poole). Seejärel Hsp 60 seob endaga 7st Hsp 15 monomeerist koosneva
kaane. Hsp 60ne kaas on oluline, kuna osaleb selles, milline valk sisse võetakse. Kui valk on
liiga suur, siis kaas ei mahu peale. Mõnedel Hsp 60ne vormidel on erinevaid kaasi (suur ja
väike). Hsp 60 suurem osa hakkab ATPd hüdrolüüsima ja hakkab oma struktuuri muutma.
Sisemised osad hakkavad liikuma, millega aktiivselt muudetakse sees oleva valgu
konformatsiooni nii kaua kuni sellel valgul ei ulatu hüdrofoobsed osad väljapoole.
Hsp 70
Töötab koos Hsp 40nega, mis on spetsiifilisuse faktor ja Hsp 70 on ATPaas. Nad seonduvad
kasvav polüpeptiidiga translatsiooni käigus, kui hakkab välja ulatuma. Nad kontrollivad peptiidi
67
ruumilise struktuuri teket. Kui on valmis sünteesitud, siis Hsp 70 ja Hsp 40 lahkuvad. Valminud
valk võib olla kas õigesti voltunud või ebakorrektselt voltunud.
Valkude lagundamine
Kui valk on oma ruumilise struktuuri kaotanud, siis ta degradeerub. Valkude lagundamine on
samuti väga täpselt kontrollitud ja suunatud protsess, mis ei toimu juhuslikult. Valgud
suunatakse lagundamisse selliste peptiidide abil, millest kõige olulisem on ubikvitiin. Ubikvitiini
kasutatakse selleks, et märkida ära neid valke, mida tuleb lagundada. Kui rakus valk on oma
struktuuri kaotanud või on valesti sünteesitud, siis sellele valgule pannakse külge ubikvitiin,
millele pannakse omakorda külge jälle ubikvtiin. Polüubikvitineeritud valk suunatakse
proteosoomi. Ubikvtiini lisamine toimub kindalt rada pidi – E1, E2 ja E3 (osalevad ubikvitiini
ülekandel märklauale). E1 seob endaga ubikvitiini, mis seotakse tioestersidemega
karboksüülotsa kaudu. see on ubikvitiini aktiveerimine. E1lt kantakse ubikvitiin üle E2le. E1 on
rakus ainult üks, E2sid on eurkarüoodi rakkudes paar tk. E3 on spetsiifilisuse faktor, mis
määrab ära milline valk läheb lagundamisele (kas hüdrofoobsed osad väljas vms). E2 küljes on
ubikvtiin tioestersidemega ja E3s lüsiini kõrval ahelaga.
????????
E3 saab mitut moodi aktiveerida. Kui valk on multiensüüm kompleksis, siis tema degradatsiooni
signaal on teiste valkudega kompleksis ja pole võimalik lagundada. Kui tuleb välja sealt
kompleksist ja ei seondud normaalsete partneritega ja signaal muutub kättesaadavaks. E3
paneb ubikvitiini otsa.
Ubikvitiini süsteem:
o Ubikvitiin on väike valk(veidi üle 100AH). Ta C-terminus on
üheahelaline – struktureerimata. Ubikvitiin on signaaliks valgu kas
transpordile või lagundamisele. Kui neid on ühe valgu küljes palju –
polüubikvineeritud – viiakse ta lagundamisele.
o E1, E2, E3 valgud osalevad selles.
o E1 – ubikvitiini aktiveeriv ensüüm. Ubikvitiin ühendatakse
tioestersidemega E1 tsüteiinisabaga.
o E2 – ubikvitiini ligaas. Suunab ubikvitiini lisamist märklaudvalkudele.
(E1 ja E2 küljes tioestersidemega, märklaual on lüsiini küljes
amiidsidemega). E1 annab ubikvitiini üle E2-le ning ise lahkub.
o Lisandub ka E3, millel on degradeeritava valgu retseptorala. E3 –
spetsiifilisuse faktor, ta tunnebki ära, milline valk kuulub
lagundamisele. Kui signaal on avatud, siis on seotud E3-ga, kui
suletud signaal, siis mitte. Kompleksis kannab E2 ubikvitiini
märklaudvalgule üle, see on juba pöördumatu reaktsioon, sest
moodustatakse peptiidside.
o Polüubikvitüleeritud valk lagundatakse.
Proteasoom
Kui valk on proteasoomis, pannakse torule korgid peale. Esimeses ringis lõigatakse ubikvitiinid
maha (proteaas, mis on võimeline amiidsidet lagundama). Proteasoomis lõigatakse valgud kuni
AHdeni. | – kui sinna satub valk, siis sulgub mõlemast otsast. Valk ei saa sealt välja enne, kui
sealt on võimelised lahkuma vabad AH-d. Kui ühte polüubikvitüleeritud valku hakatakse
68
lagundama, siis ikka lõpuni. Proteasoomi suunatakse valgul vastavalt N-terminaalsele
aminohappele, mis on indikaatoriks ka tema vanuse suhtes. Õige AH on esimesena alati ju
metioniin. Tekkinud N-terminus määrab ära valgu eluea.
On AH-d, mis destabiliseerivad valgu, kui üks nendest on N-terminaalne, siis on eluiga lühike.
N-otsa reegel
•
Erineb veidi inimesel (valgu eluiga tundides või päevades) pärmil ja bakteril
(valgu eluiga minutgites kuni tundides)
•
(Asn ja Asp bakteris stabiliseeriv, euk destabiliseeriv)
•
Cys inimeses destabiliseeriv, pärmis, bakteris stabiliseeriv
Valkude eluiga on kontrollitud N-terminaalse AH poolt. See AH, mis on valgus esimene.
Metioniin enamik juhtudel lõigatakse ko-translatsiooniliselt ära juba translatsiooni käigus. Kui
valgu N-terminuses on 1 10st AHst (Arg, Lys, His, Phe, Leu, Trp, Ile, Asp, Glu, Asn, Gln), on
signaaliks polüubikvitinüleerimisele ja suunamisel proteasoomi. Nende eluiga on lühike (10-20
min). Kui on ülejäänud 10 AHd, neid nii kiiresti ei lagundata (lagundamise signaal pole N-
terminuses), mida kutsutakse destabiliseerivateks AHdeks.
Inimese, loomade valkude eluiga on suhteliselt pikk (isegi vähem stabiilsed võivad elada tunni).
Vabalt elavatel mikroorganismidel on eluiga palju lühem (minutid kuni tunnid). Asn ja Asp on
bakterites stabiliseeriv aga eukarüootides destabiliseeriv. Cys vastupidine???
ClpP – chaperon like protein. Proteaas, mis töötab koos chaperonidega (kui chaperon ei suuda
valgu struktuuri õigeks teha). Moodustab toru.
Persistorid ? Üks antibiootikumi meetod bakteri tapmisel:
o Paljud inhibeerivad valkude sünteesi. Kui süntees jääb pooleli, siis
tekivad ju poolikud valgud, valgufragmendid ja hakatakse
sünteesima hulgaliselt proteaase. Lõpuks hakkavad proteaasid
lagundama ka mitte lagundamist vajavaid valke. Lõpuks rakk
hukkub, valke pole.
o ClpP valk – chaperone like protein P. On ühelt pootl chaperone,
teisalt proteaas. Moodustab proteasoomi sarnaseid struktuure, ilma
punnideta otstes.
o Tal on suletud vorm, mis ei tee miskit.
o Avatud vorm lagundab valgu aminohapeteks.
o On leitud ühend atsüülpepsipetiid, mis aktiveerib ClpP valku. On
olemas persistervalgud, mida selline antibiots muidu ei tapa, neil ple
aktiivset valgusünteesi.
o AGA kui lisada atsüülpepsipeptiidi(ADEP), siis tapetakse ka
persisterid.
o EHK võimendab antibiootikumi võimet-toimet oluliselt!
Hiire fibroplastide kultuur
Keskmine mRNA eluiga on 9 h, valgu eluiga 46 h. Ühte mRNAd raku kohta on keskmiselt 17
koopiat ja valku 16 000. Seega sünteesitakse 40 valgumolekuli ühelt mRNAlt tunnis. mRNA
koopiate arv on seotud valgu koopiate arvuga – mida rohkem mRNAd on rakus, seda rohkem on
vastavat valku (statistiline seos). mRNA tase rakus ei määra üheselt valgu taset rakus, kuid
69
määrab kui palju mingit valku on (ei ole ühene funktsionaalne seos). Erinevaid valke
sünteesitakse väga erineva kiirusega (erineb kordades). Suurim kiirus on 180 koopiat tunnis.
Valke sünteesitakse erineva kiirusega seetõttu, et valgud on erineva suurusega. Väiksemat
sünteesitakse kiiremini kui suurt. Keskmiselt ühe mRNA kohta tehakse 450 valgumolekuli. 20%
valkude sünteesiks kulub 80% valgusünteesile kuluvast energiast, 80% ribosoome on hõivatud
20% mRNAga. Need geenid, mis ekspresseeruvad madalal tasemel, neil ei ole vahet
missuguseid koodoneid kasutatakse. Vähearvukate valkude eluiga ei mõjuta oluliselt raku
energiamajandust (kui on vähetähtis).
Globaalne geeniekspressiooni regulatsioon
mRNA lagundamise kiirus määrab alla 10% kogu valgu tasemest. Valgu sünteesi kiirus on kõige
olulisem parameeter valgu koguse määramisel rakus. Valgu lagundamine on suhteliselt vähe
oluline.
Kui vaatame korraga vga paljusid valke, siis näeme et transkriptsiooni tasemel regulatsiooni ja
translatsiooni tasemel regulatsiooni kiirus on võrdselt olulised geenide avaldumise
kontrollimisel. Teistel tasemetel on väiksem tähtsus.
Epigeneetiline pärilikkus ja geeni vaigistamine
Epigeneetiline nähtus – need regulatsioonimehhanismid, mis asuvad geenide peal.
19. sajandi arengubioloogid jagunesid epigeneetikuteks ja predeterministideks. 20 sajand oli
geneetikute (predeterministide) aeg
Fenomenid:
Post-transkriptsiooniline geeni vaigistamine – sünteesitakse RNA, aga selle pealt valku
ei sünteesita või sünteesitakse mittefunktsionaalne valk
Pärilik DNA modifitseerimise muster – need geenid, mille järjestus võib olla täpselt
ühesugune kahel inimesel, aga nende geenide modifitseerimise muster (metüleerimine
eelkõige) on erinevad ja seetõttu ühel see geen avaldub ja teisel mitte (nagu poleks seda
geeni)
Päritav histoonide koht-spetsiifiline modifitseerimine – histoonid H3 ja H4 jäävad DNAga
seotuks sama DNA piirkonnaga, H2A ja H2B dissotseeruvad. Kuidas H3 või H4 N-terminaalne
saba on modifitseeritud
Priioni nähtus (valguline pärilikkus)
Hsp 90 osa vaikivate mutatsioonide kogumisel
70
Post-transkriptsiooniline geeni vaigistamine
mRNAga komplementaarne RNA on antisense.
Fire ja Mello: võtsid C. elegans’i ja lisasid talle mRNA tüki, mis oli samasuunaline ja sama
järjestusega nagu ussikese lihasvalku kodeeriva mRNA järjestus või antisense järjestuse. Midagi
71
ei juhtunud, ussike elas edasi. Kui senes ja antisense kokku said ja tekitada sellest
kaheahelaline mRNA ja lisada ussikesele, siis ussike hakkab tõmblema ja käitub nii nagu siis kui
lihasvalku pole. Lihasvalgu tase langeb. Kaheahelaline RNA blokeerib valgu sünteesi.
Kaheahelaline RNA läheb rakku. Rakus lõigatakse tükkidest, mis lahutatakse eraldi ahelateks.
Tükike seondub valgulise kompleksiga rakus, mis kasutab seda järjestust, et otsida välja RNAga
komplementaarne mRNA ja suunab mRNA lagundamist. Kui toimub paardumine, siis mRNa
lagundatakse.
Transkriptsiooni vaigistamine – ei toimu transkriptsiooni, ei sünteesita RNAd. Geeniprodukte ei
akumuleeru, sest ei toimu transkriptsiooni, näit. kromatiini struktuuri tasemel vaigistamine
Transkptsiooni järgne vaigistamine – ei sünteesita funktsionaalset valku, kuid RNA
sünteesitakse. Toimub geeni transkriptsioon, kuid valku ei akumuleeru, sest mRNA
lagundatakses või on häiritud valgu süntees
RNA interferents e RNAi – Kaheahelalise RNA poolt käivitatav protsess, mille käigus
komplementaarne mRNA lõigatakse ja lagundatakse.
Nähtus, mille käivitab kaheahelalise RNA sattumine rakku. See mRNA mis on komplementaarne
ühega nendest kahest ahelast, see mRNA lagundatakse. Väike RNA (siRNA) segab vahele
mRNA molekuli stabiilsusele. Avastasid Fire ja Mello. Kaheahelaline RNA lõigatakse tükkideks,
mis on 21-25 nukleotiidi paari pikad. Kummaski otsas 2 nukleotiidid, mis pole paardunud (3’
üleulatuvad otsad). Hakati otsima geeni eukarüoodis, mis oleks sarnane bakteri RNaas 3ga.
Leiti produkt, mille nimeks pandid dicer. Sarnase ruumilise struktuuriga, tunneb ära
kaheahelalise RNA ja lõikab kaheahelalise RNA lühimateks tükkideks. Edasi need
kaheahelalised RNA tükid lähevad rakus kompleksid, mille nimi on RISC. Kaheahelalised tükid
lahutatakse üheahelalisteks. RISC jääb seotuks lõpuks üheahelalisega. ??? Argonaute 2 lõikab
RNA katki.
siRNA
(small interfering)
21–25 nt, pakitakse koos valkudega RISC (RNA-induced silencing complex) kompleksi.
siRNAd omavad täpset komplementaarsust märklaud RNA-dele ja indutseerivad RNA-
interferentsi s.o märklaudgeeni lõikamist ja degradatsiooni.
miRNA (microRNA)
Raku poolt kodeeritud rakusisene, mis on enam-vähem sama pikad kui siRNA. On ka
kaheahelalised. Sageli pole ahelad täies ulatuses komplementaarsed. 3’ otsas on 2 ülaulatuvat
nukleotiidi, mis on põhjustatud asjaolust, et ta on sama ensüümi produkt nagu siRNA. On pärit
rakutuumas olevatelt geenidelt, RNA polümeraas II produktid. Sisaldavad sellised RNA
järjestusi, mis moodustavad juuksenõela struktuure. Dicer lõikab välja kaheahelalised väiksed
minad, mis seostuvad ka RISCiga. miRNA võidakse lagundada või ka mitte. Translatsiooni
pidurdatakse alati. miRNA suunab mRNA deadenüleeritakse, cap võetakse ära ja siis
lagundatakse????. Võib ka suunata translatsiooni inhibeerimisse?????. miRNA puhul on oluline,
et mRNA lõigatakse katki ja suunatakse lagundamisse.
miRNA puhul ei teki perfektne komplementaarne ahel mRNAga. Pooled nukleotiidid on
komplementaarsed ja pooled ei ole. Kui mRNAd hakatakse lagundama miRNA sõltuvalt, siis
RISC kompleks suunab kõigepealt mRNA deadenüleerimist 3’ otsast (eemaldatakse polü-A
72
saba) ja 5’ otsast võetakse ära m7G (cap). Sellega mRNA kaotab oma identiteedi, pole enam
mRNA.
mRNA stabiilsuse determinandid on need, mis määravad kui kaua mRNA elab. Need
determinandid jäävad stopp-koodoni ja polü-A vahele. Kui sellega seondub miRNa, siis suunab
sellega polü-A lühendamist.
miRNA on geeniekspressiooni regulaator. Üks miRNA reguleerib väga paljude geenide
aktiivsuse taset.
miRNAde biogenees
Paljud on pärit intronitest. Intronis moodustuvad lassostruktuurid, mis reeglina lagundatakse,
aga nad võivad sisaldada ka funktsionaalseid geene. Paljud nendest geenidest, mida intronid
sisaldavad, on just RNA tasemel toimivad regulaatormolekulid. Intron moodustab kaheahelalisi
struktuure, millest lõigatakse välja küps miRNA, mis transporditakse RISC kompleksi, kus
tehakse ta üheahelaliseks. miRNA, mis on pärit intronist, võib reguleerida sama geeni.
Ei pea alati pärit olema intronitest, vaid neil on ka omad geenid (miRNA geenid). Tüüpilise
geenid, mis kodeerivad miRNAsid. Primaarsest transkriptist, mis moodustab kaheahelalisi
struktuure, tekib pri-miRNA. miRNAd tuleb kõigepealt lahutada, mida viivad läbi ensüümid
Drosha/Pasha. Eksportiin 5 transpordib pri-miRNA tstüplasmasse, Dicer lõikab õige pikkusega
miRNA. miRNA läheb RISC kompleksi, kus tehakse ta üheahelaliseks ja suunab translatsiooni
repressiooni. RISC kompleks koos mRNAga, mis on miRNA järjestuse lausel ülesehitatud,
pakitatakse tihedalt sisse ja säilitatakse P-kehades. Võib sealt võtta uuesti kasutusele kui vaja
(mRNA säilitamine).
P-kehad
miRNA võib suunata mRNA lagundamist, aga igal juhul kipub ta suunama translatsiooni
vaigistamist. Vaigistatav mRNA suunatakse P-kehasse.
P-kehad on seotud RNA metabolismiga. Mitmesugused RNAga seotud valgud ja RNA molekulid
on pakitud P-kehakesse, mis on väiksed ja tihedad. U 1910 avastati.
RNA interferents (RNAi)
RNA interferents on eriti oluline taimedel. Enamik taimeviirustest on RNA viirused. Taimed
kasutavad nende viiruste vastu võitlemiseks RNAi-d. Toimub üle siRNAde. Kui inimesel on 1
Dicer geeni juures, taimedel on 6 erinevat Dicerit. Mitte ainul taimedes vaid ka mõnedes
loomades on leitud, et interferentsi nähtus võib levida ühelt rakust teisele. Tekkinud märklaud
RNA tükkidest (kaheahelalised, 22-24 nukleotiidid pikad) RdRP sünteesib RNAle teise ahela
juurde ja need väiksed RNAd transporditakse järgmisse rakku. Transport vesiikulite abil
(miRNAd või siRNAd sees). Vesiikulid ühinevad märklaudrakkudega. miRNAd hakkavad
reguleerima märklaud rakkude geeni ekspressiooni. Võivad tekkida sekundaarsed siRNAd, mis
muudavad immuunseks primaarsed siRNAd ???
Rakku tunginud kaheahelaline RNA lõigatakse juppideks, mis lähevad RISC kompleksi. Argonaut
lahutab kaheahelalise RNA, otsib märklaua ja lagundab RNA. Nendest tekkivatest RNAde
tükkidest võidakse sünteesida uusi RNAsid – sekundaarsed siRNAd.
73
miRNAd võivad indutseerida mRNA lagundamist
mRNA algselt kaheahelalised, komplementaarsed märklaud RNAga, toimivad kompleksis RISC.
Kõige tähtsam komponent RISC süsteemis peale mRNA on valk argonaut. Ristkompleksis
võidakse Mrna pakkida P kehadesse (viia translatsioonist välja) või lagundatakse. Lagundamine
ei toimu kohe kui mi RNA paardub vaid lagunemisele peab eelnema mRNA märkide
mahavõtmine (5’ otsas tuleb eemaldada CaP ja 3’ otsast polü A saba). Seejärl lõigatakse mRNA
katki mille tulemusel on ta substraadiks sellisele struktuurile nagu epsosoom.
mRNA stabiilsuse reguleerimist piirab koht kuhu mRNAd seounduvad, on 3’ mittetransleeritav
ala, kus on miRNA äratundmiskohad. miRNA osad seonduvad selles piirkonnas olevate
järjestustega. Üks miRNA reguleerib väga paljude geenide aktiivsust.
RNA interferents (RNAi)
Taimede viiruste kaitsemehhanism- RNA interferentsimehhanism. Taimeriigis eriti palju levinud.
See RNA interferents toimub üle siRNAde. Mis pärineb rakku tunginud RNAst nt viiruse RNAst.
Rakku tunginud kaheahelalisest DNA lõigatakse välja jupid, mis on 24 nukleotiidi pikad, mis
ähevad ristkompleksi, seal otsitakse siRNAle komplementaarne ahel üles ja RNA lagundatakse.
See protsess võib edasi minna nii, et lagundatud RNAdest tekivad tükid, millest sünteesitakse
uued väikesed RNAd- sekundaarsed siRNAd (alguses rakku tunginud on primaarsed siRNAd)
mis levivad juba naaberrakkudesse. Kui naaberrakus on siRNA juba olemas siis tal pole
võõimalust paljuneda, sest siRNAde abil viirus lagundatakse. Ei toimu samas rakus vaid ka
naaberrakkudes.
RNA interterferents mõjub ka transkriptsioonile. Vaigistamisel tekkinud siRNAd lähevad mitte
ristkompleksi vaid RICS kompleksi- RNA indutseeritud transkriptsiooni vaigistamise kompleks-
seonduvad argonaut1 ga. Selle vahendusel seondub DNAga (kromatiiniga) ja suunab kromatiini
metüleerimist ja geeni vaigistamist. Nt histoonide metüleerimine võib viia geeni vaigistamisele.
siRNA vaigistamise mehanism
siRNA paardub mRNAga, argonaut on tähtsaim komponent. Argonaut lagundab mRNA ja enne
seda suunab, et see üles leida. Argonaudil on nukleaasne aktiivsus. siRNA poolt indutseeritud
vastus võib levida ka kromatiini tasemele.
Argonaut- RNA helikaasne aktiivsus, lahutab miRNA ja siRNA ahelad, valib sealt ühe ahela. Ühe
ahela alusel otsitakse mRNA, millega miRNA on komplementaarne ja argonaut võib lõigata
katki mRNA. Kui mRNAd ei lõigata siis RNA viiakse P kehadesse ja pakitakse partiklitesse kus
RNA ei ole translatsioonimehanismile kättesaadavad.
miRNAd esinevad igal pool ja miRNA sarnased esinevad ka bakterites. miRNAde
väljalõikamine eellastest on erinev erinevates organismides.
Taimedes: on meil esimene protsess- põhiline ensüüm on Dci ja erinevad Dci protsessivad
miRNA valmis, valmis protsessitud kaheahelaline miRNA transporditakse tsütoplasmasse, kus
läheb RISC kompleksi
loomad: miRNA eellane pre-miRNA mille lõikavad välja Drosha ja Pasha. Tsütoplasmasse
transporditakse mRNA eellane pre-miRNA ( pikk üheahelaline ling mis on iseendaga
komplementaarne). Lõplik kujunemine toimub tsütoplasmas, kus üks ahel jääb argonaudiga
seotuks ja teine lagundatakse
74
Kaheahelaline miRna koos agronaudiga, ahelad lagundatakse, valitakse välja efektorahel, teine
ahel lagundatakse. Teine etapp võtab aega- märklaud RNA ülesotsimine.
RISC kompleks
Tähtsaim komponent on argonaut, mida on selles kompleksis vähemalt 2 (inimesel 6). Argonaut
(Ago) koosneb mitmest RNA-siduvast domeenist, millest PAZ domeen tunneb ära äikese RNA3’
otsa, PIWI (osaleb samuti geenivaigistamises) domeen seondub nii miRNA kui märklaud RNAga.
Ago valib välja toimiva miRNA ahela 5’ otsa alusel, lahutab RNA ahelad ja otsib välja
komplementaarse mRNA märklaua. Ago II on ka nukleaas, mil lagundab märklaud RNA
siRNA pole ainus kompleks mis suunab geenivaigistamist, mõned telomeeride piirkonnas on
selleks oma süsteem- spetsiifilised histoonide deatsetülaasid (sir 3 ja sir 4) mis suunavad
histoonide deatsetüleerimist ja seetõttu telomeeride tihedas kokkupakitud osas ei saa geenid
ekspresseeruda
RNAi kasutamise meetod.
Hästi võimas uurimise meetod. Bakterite raamatukogu- koht kus erinevad kloonid toodavad
erinevaid pikki kaheahelalisi RNA-sid, lisatakse toidule. Drosophila rakud võtavad RNA söötmest
ise sisse. Organismi tasemel- süstimine embrüodesse. Transfektsioon või nakatamine
retroviirusvektorite raamatukoguga, mis sisaldavad erinevaid shRNAsid. Mõnede ussikestega
pärast seda, läheb elu viletsamaks- põhjustas ussigenotüübi muutuse. Saame geenid välja
lülitada. Saame geenide funktsiooni uurida ilma et oleks muutunud nende struktuuri. Imetajate
puhul võib seda teha valmis tehtud kaheahelaliste siRNAdega, millega saab kindlaid geene
välja lülitada või siis retro viirus vektoreid.
Transgeensed taimed
Võimalik teha viiruskindlaid taimi, mida viirused enam ei nakata. Kui me viime kartulisse sisse
väikese DNA mis on kartuliviirus A genoomi osa, et kartulist sünteesitakse kartuliviirus A RNAd
mis on osa kartuliviirus A genoomist, siis me indutseerime nendes taimedes RNA interferentsi
ja kartul muutub resistentseks. Taimed mis on resistentsed putukate kahjustuste vastu.
Tehakse taimed mis on putukatele nii kibedad, et putukas ei taha seda süüa. Seemneteta
tomatid- kalkooni süntaas (aine mis on seemnekesta põhikomponent) saame välja lülitada RNA
interferentsiga kalkooni süntaasi. Nii saame valida mis tomatitel on seemned ja millistel mitte
RNAa
RNA aktiveerimine. Mitmete tuumor suppressor geenide puhul on nähtud RNAi’le vastupidist
efekti, siRNA rakku viimine tõstab märklaudjärjestusega geeni avaldumise taset.
Üks siRNA võib vaigistada üldse mingit teist geeni mis on mingi geeni regulaator aktivaator.
Lisaks miRNAdele on eriline klass RNAsi piRNA
piRNA- kohe algusest peale üheahelalised, et mooduste kaheahelalisi juuksenõela struktuure.
piRNAd seonduvad kõik valguga PIWi. Igal piRNAl on oma pisike geen. PIWI ON ARGONAUDI
TÄDIPOEG!!!!!! PIWIgasuunab RNA välja lülitamist. Funktsioneerivad transposoonide
vaigistamisel. Kontrollib oma peremehe geenide ekspressiooni ja vaigistab transposoone.
Epigeneetiline pärilikkus- selline pärilikkus mis ei sõltu otseselt DNA järjestusest, järjestus
sama aga tulemus erinev
75
Muul ja Hinni
Muul on eeslitäku ja hobusemära järglane. Emasid muulasi kutsutakse mollideks, sellest pooled
on steriilsed. Isad on kõik steriilsed. Kui Hobusetäkku paaritada eeslimäraga- saame hinni.
Muula ja hinni geenid on ühesugused, geenid on samad aga neil on võrdlemisi erinev
välimus.Kõik hea on läinud muulale ja kõik halb on hinnil. Muulale mahub palju väikseid
kauboisid peale aga hinnist asja ei saa.
Mesilased
Emamesilane erineb töömesilasest ainult geenide metüleerimise poolest.
X kromosoomi vaigistamine Xist RNA abil
Xist RNA geen on X kromosoomis ja selle avaldumine lülitab kromosoomi välja in cis
CRISPR/Cas süsteem, geenide vaigistamine bakterites
Bakterid suudavad sama meetoditega võidelda nii RNA kui DNA viiruse vastu. Väikesed RNA
kordused, mis sünteesitakse lähevad genoomis kindlasse piirkonda, kus nad on bakteri geeni
regulatsiooni kontrolli all. Väikesed RNAd on suutelised suunama märklaud RNA ja märklaud
DNA lagundamist.
DNA vermimine isa ja emaliinis
Olenevalt kas geen on pärit emalt või isalt, on ka geeni avaldumine erinev.
Eluajal omandatud modifitseerimise muster võib olla päritav (tagab säilitav metülaas). Kui
vanem ahel on metüleeritud ja uus ahel pole metüleeritud, siis säilitav metülaas metüleerib
tütarahela kasutades vanemahela metüleerimise mustrit
Prion
Valgu ruumilisel struktuuril põhinev pärilikkus. Erineb teistest pärilikkuse vormidest, sest on
suhteliselt vähe soetud DNA järjestusega. Ta on küll DNA järjestuse alusel sünteesitud, kuid
priini pärilikkus sõltub valgu ruumilisest struktuurist, võib mõjutada teiste valkude ruumilist
struktuuri. Priion valkudel võib olla mitu erinevate ruumilist struktuuri.
Priion esineb hulkraksetel, aga ka pärmidel, annab kohastumusliku eelise. Inimese jaoks on
oluline, sest prion on soetud mitmete neurodegeneratiivsete haigustega, amüloidide tekkega
kesknärvisüsteemis. Muudavad aju käsna taoliseks, lõpuks ei jää seal enam neuronitele erilist
ruumi. Prion põhjustab veiste käsnataoline entsefalopaatia ehk hullulehmatõbi. Selle
tulemusena Uks hukati sadu tuhandeid veiseid. Tema nakkuslikkus on erakordselt haruldane.
Ravida ei osata. Kandub üle inimesele, kutsutakse
Creutzfeldti-Jakobi tõbi, süües haige lehma
kesknärvisüsteemi, väga väikese sagedusega. Britid toitsid veiseid veisekondi jahuga, sealt
leviski see haigus. Lammastel esinev kratsimistõbi kandub ka inimesetele üle,
kesknärvisüsteemi haigus, on põhjustatud sama valgu poolt. Kuru haigus kirjeldatud ainult
Paapua Uus-Guineas ja seostatakse kannibalismiga, aju just.
Prion valk ekspresseerub neurogliia rakkudest, on seotud mälu funktsiooniga. Prioni domään
esineb kahes konfirmatsioonis. Üks on täies ulatuses alfa heeliksitest koosnev struktuur, see
76
võib üle minna selliseks struktuuriks, mis koosneb põhiliselt beeta heeliksitest, mõned alfa
lehed jäävad alles ka. See üleminek võib toimuda iseenesest, aga toimub väga-väga harva.
Beeta struktuur nakatab alfa vormi niimoodi, et ka teine prioni molekul muutub beeta
struktuuriks. Beeta struktuurid kleepuvad üksteise külge ja moodustavad amüloide, mis on
lahustumatud ja neid enam tagasi ei saa muuta. Kui on üks beeta vormis prion, pole teistega
kokku kleepunud, siis on veel väike võimalus sellel tagas muutuda. Beeta struktuur tekitab
patoloogiat.
C. H. Waddington epigeneetiline maastik: organim hakkab arenema, siis tal on palju erinevaid
võimalusi, mis suunas minna, aga tagasi enam ei saa, kui on ühe suuna valinud. Geneetilist
mitmekesisust maskeerib Hsp 90.
S. Lindquist hakkas uurima Hsp90 osa mutatsioonide avaldumisel. Leidis, et kui organismis on
olemas funktsionaalne hsp90, siis enamus Drosophia mutatsioone ei avaldu fenotüübiliselt. Kui
hsp90 inaktiveerida, siis avalduvad mutatsioonid. See käib samuti valgu ruumilise struktuuri
kaudu. hsp90 on heat shock proteiin, chaperon, mõjutab teiste valkude konfirmatsiooni, kui ta
ei suuda teisi valke õigeks konfirmatsiooniks väänata, siis saadab ta lagundamisele.
Geenid ja evolutsioon
Evolutsiooniliselt kaugetel genoomidel on siiski lähedasel arvul geene:
Nematood C. elegans 20 060
Äädikakärbes 14 000
Meritupp 15 000
Üks asi on see, kui palju on geene, aga teine asi on kui palju on DNAd raku kohta.
Kõige väiksemad genoomid on endosümbiontsetel bakteritel:
•
Carsonella ruddii 160 kb
•
Buchnera aphidicola 422 kb
•
Osa geene on “ümber kolinud” peremehe kromosoomidesse.
On selliseid endosümbiontseid baktereid, kes elavad teise raku sees, paljunevad ainult teise
rakuga koos, väljaspool peremeest ei esine.
DNA sisaldus raku kohta ei tähenda, et neil oleks rohkem geene. Erinevate valkude arv ei
tulene otseselt geenide arvust. Kõige suuremad genoomid on salamandritel ja liiliatel, umbes
100x suurem kui inimesel. Kui inimese DNA hulk raku kohta on 3*109 aluspaari, siis nii
salamandritel kui ka liiliatel võib olla see DNA hulk raku kohta olla 1011.
Genoomi suurus: DNA sisaldus raku kohta ei tähenda, et neil oleks rohkem geene.
Erinevate valkude arv ei tulene otseselt geenide arvust.
Mudel-liigid
Nende geenide alusel tehti järeldusi kõikide teiste organismide järgi. Tänapäeval veidi
teistmoodi. On palju organisme, kelle kohta teatakse ainult DNA järjestust, aga ei teata
välimus jm.
77
Geeni puu
Geene tekib ainult teistest geenidest. Igal geenil on eellane. Geenid võivad muutuda ja
evolutsioneeruda, võivad omandada uue funktsiooni. Ühe geeni järglased moodustavad
geeniperekonna.
Kõik GTPaasid on omavahel suguluses, nad on pärit ühest algsest geenist. Nende
ruumiline struktuur on väga sarnane, AHline järjestud ei pruugi samas üldsegi sama
olla. . ATPaase on aga väga palju erinevaid ehk mitteseotud perekondi.
Geen on DNA või RNA lõik, mis kodeerib ühte eralduvat produkti. Looduses iseseisvalt DNA
sünteesi ei toimu, RNA süntees võib küll toimuda, pole vaja matriitsi ette RNA sünteesil.
Ilma juureta geeni puu – fibroplasti kasvufaktori geenid. Tekkinud ühisest eellasest. AHde
asendumise käigus on valgud omandanud uue funktsiooni ja tekkinud täiesti uus funktsioon.
Geeni puu – ei tea milline on kõige vanem. Järjestuse alusel grupeeritud. Näha ka kui palju
asendusi on
Uued geenid moodustuvad duplikatsioonide tulemusena. Üks täidab oma ülesannet edasi, teine
hakkab muutuma. Beeta-globiinide on heaks näiteks. Järjestuse sarnasuse alusel saab nende
globiinide geenide suguluse ritta panna. Nt beeta-globiini geenide evolutsioon. Haikalal on 1
beeta-globiin, inimesel aga mitu. Inimese erinevad beeta-globiinid erinevad omavahel sama
palju kui üks haikala ja inimese globiin (valgu poolt erinevus).
Mutatsioonid
Mutatsioonid enamasti erilist fenotüübilist efekti ei põhjusta. Enamustel mutatsioonidel on efekt
valgu struktuurile väga väike, selle mutatsiooni esinemise tõenäosus on suurem, kui suurt
efekti põhjustavatel.
Mutatsiooniga saab muutuda valgu järjestus või ka märklaud, millega see valk seostub. Kui
transkriptsiooni faktoris muutub mutatsioon, siis ta võib hakata ära tundma teistsugust
geenijärjestust – muutub geeniregulatsioon. Mutatsioon võib aga tekkida nii, et see
transkriptsiooni faktor hakkab ära tundma hoopis teistsuguseid geene, mida ta ennem ära ei
tundnud.
Oluline ei ole ainult, milline geen ekspresseerub, vaid ka kuskohas ta ekspresseerub.
Drosophila embrüos jäävad tütarrakud seotuks torudega (ühendustega). Geenid, mis
avalduvad ühes rakus, saavad liikuda kõikidesse rakkudes. Hulk morfogeene, mis
määravad äädikakärbse arengut, embrüos ei avaldu aga emarakus olemas. Drosphila
embrüos tekivad erinevate organite imaginaardiskid, rakkude rühmad, mis määravad
mingisuguse organi arengu. Lülijalgsete embrüo koosneb segmentidest, kus erinevates
segmentides avaldub erinev komplekt geene. Oluline on, et kus kohas mingi geen
ekspresseerub (kus ja milline). Kui panna silmaarengut määrav geen jala
imaginaardisikile, on neil jalgadel silmad.
Hox geenid on organismide arengus väga tähtsad, määravad teiste geenide avaldumise mustri.
Mutatsioonid
FOXP2 geenis on seotud kõnevõimega, neid on nii inimesel, lindudel, hiires,
šimpansil. Inimese geen erineb teiste sugulaste geenides vähe. Kui hiirel asendati FOXP2
geenivariant inimese geeniga, siis muutisid hiired nutikamateks. FOXP2 avaldub ajus ja teistes
78
kudedes. See geen on häälitsemisefaktorina tuntud. Ta on transkriptsioonifaktor, mis reguleerib
300-400 geeni nii embrüos kui täiskasvanud organismis. Lindudel mõjutavad FOXP2 geeni osad
lindude võimet laule õppida.
Inimlikkuse geen
DNA piirkond, mis inimesel erineb kõige rohkem teistest primaatidest see kodeeriv RNAd, see
on
HAR1 regioon. Kõigil inimese sugulastel on see kontsentreerunud, väga sarnane, kuid
inimsesel endal on see hoopis teistsugune, inimesel on seal 18 mutatsiooni võrreldes
šimpansiga, väga palju. šimpansil ja gorilla näiteks on seal keskeltläbi 1 mutatsiooon ainult. See
piirkond kodeerib ühte väikest RNAd, väike RNA võib mõjutada väga pajude geenide aktiivsust.
HAR1F kodeeritud RNA avaldub neokorteksis ehk migreeruvates neuronites, nendes neuronites,
mis moodustavad ajukoore.
RNA........................................................................................................................ 1
Nukleiinhapete struktuur........................................................................................2
DNA/RNA suur ja väike vagu...................................................................................3
DNA kaksikheeliks.................................................................................................. 4
DNA topoloogia....................................................................................................... 5
Kromatiini struktuur................................................................................................ 6
Nukleosoom............................................................................................................ 6
DNA replikatsioon................................................................................................... 7
DNA otste replitseerimine eukarüootides.............................................................10
DNA replikatsioon raku kontekstis........................................................................10
Replikoni mudel.................................................................................................... 11
Initsiaatori funktsioonid replikatsiooni alustamisel...............................................11
79
DNA replikatsiooni alustamine päristuumsetes....................................................12
Kromosoomi otste replikatsioon: inimesel............................................................13
DNA mutatsioonid ja nende parandamine............................................................13
MutS valk, ahela lagundamine.............................................................................14
Restrikitsioonid-modifikatsioonid..........................................................................15
Modifikatsiooni-restriktsiooni süsteemid bakterites:.............................................15
Reparatsioon........................................................................................................ 15
DNA reparatsiooni mehhanismid..........................................................................16
DNA reparatsiooni süsteemid: (tüüp-viga-ensüümid)...........................................17
NVR reaktsioonid – Uvr teema..............................................................................17
RNA reparatsioon................................................................................................. 18
Valkude parandamine.......................................................................................... 18
Homoloogiline rekombinatsioon:..........................................................................18
Transposoonid...................................................................................................... 22
T-DNA: bakterist üle kantav DNA..........................................................................26
Transkriptsioon..................................................................................................... 27
RNA polümeraas................................................................................................... 29
Promootor............................................................................................................. 29
Erinevad geenid.................................................................................................... 30
Prokarüootidel transkriptsiooni regulatsioon........................................................31
Transkriptsiooni initsiatsioon, regulatsioon..........................................................31
Kromatiini struktuur.............................................................................................. 40
Histoonide modifikatsioonide kaks toimet............................................................42
Histooni H3 kovalentsed modifikatsioonid omavad erinevat tähendust...............42
Eukarüootide pre-mRNA splaissimine - kokkulõikus.............................................44
Intron-ekson skeem.............................................................................................. 44
RNA splaissimine.................................................................................................. 44
ATAC intronid ehk minoorsed intronid..................................................................46
Kotranskriptsioonilised sündmused pre-mRNA sünteesil......................................48
Valgu biosüntees.................................................................................................. 49
Universaalne geneetiline kood.............................................................................49
Lugemisraam ORF (avatud lugemisraam)............................................................51
Koodonite kallutatus............................................................................................. 52
CAI – koodonite adaptatsiooni indeks...................................................................52
Standardiseerimine.............................................................................................. 53
Koodonite kontekst............................................................................................... 53
DNA ja RNA koostis............................................................................................... 53
80
tRNA sekundaarstruktuur.....................................................................................54
tRNA identsus....................................................................................................... 55
Translatsioon........................................................................................................ 55
Initsiatsioon prokarüootides.................................................................................56
Initsiatsioon eukarüootides...................................................................................56
Elongatsioon......................................................................................................... 57
Terminatsioon....................................................................................................... 57
Ribosoomi struktuur............................................................................................. 58
Ribosoomi komponendid......................................................................................58
Antibiootikumid ja valgusüntees..........................................................................61
Antibiootikumide resistentsuse tagavad erinevad teed........................................62
Mutatsioonid ribosoomides tagavad resistentsuse ABde suhtes..........................62
tmRNA.................................................................................................................. 63
Bakterite ja eukarüootide translatsiooni võrdlus..................................................63
RNA kvaliteedi kontroll päristuumsetes................................................................64
Nonsense mediated decay...................................................................................64
Translatsiooniline kontroll....................................................................................65
No-go decay......................................................................................................... 65
Transkriptsiooni aktivaator GCN4 norm rakus......................................................65
Ko-translatsiooniline transport.............................................................................66
Valgu voltumine................................................................................................... 66
Chaperon.............................................................................................................. 67
Hsp 60 suunatud voltumine..................................................................................67
Hsp 70.................................................................................................................. 67
Valkude lagundamine........................................................................................... 68
Proteasoom.......................................................................................................... 68
N-otsa reegel........................................................................................................ 69
Hiire fibroplastide kultuur.....................................................................................69
Globaalne geeniekspressiooni regulatsioon.........................................................70
Epigeneetiline pärilikkus ja geeni vaigistamine....................................................70
Post-transkriptsiooniline geeni vaigistamine........................................................71
miRNA (microRNA)............................................................................................... 72
miRNAde biogenees............................................................................................. 73
P-kehad................................................................................................................ 73
RNA interferents (RNAi)........................................................................................73
miRNAd võivad indutseerida mRNA lagundamist.................................................74
RNA interferents (RNAi)........................................................................................74
81
siRNA vaigistamise mehanism..............................................................................74
RISC kompleks...................................................................................................... 75
RNAi kasutamise meetod.....................................................................................75
Transgeensed taimed........................................................................................... 75
RNAa.................................................................................................................... 75
Muul ja Hinni......................................................................................................... 76
Mesilased.............................................................................................................. 76
X kromosoomi vaigistamine Xist RNA abil............................................................76
CRISPR/Cas süsteem, geenide vaigistamine bakterites........................................76
DNA vermimine isa ja emaliinis............................................................................76
Prion..................................................................................................................... 76
Geenid ja evolutsioon........................................................................................... 77
Mudel-liigid........................................................................................................... 77
Geeni puu............................................................................................................. 78
Inimlikkuse geen.................................................................................................. 79
82
83
Document Outline
- RNA
- Nukleiinhapete struktuur
- DNA/RNA suur ja väike vagu
- DNA kaksikheeliks
- DNA topoloogia
- Kromatiini struktuur
- Nukleosoom
- DNA replikatsioon
- DNA otste replitseerimine eukarüootides
- DNA replikatsioon raku kontekstis
- Replikoni mudel
- Initsiaatori funktsioonid replikatsiooni alustamisel
- DNA replikatsiooni alustamine päristuumsetes
- Kromosoomi otste replikatsioon: inimesel
- DNA mutatsioonid ja nende parandamine
- MutS valk, ahela lagundamine
- Restrikitsioonid-modifikatsioonid
- Modifikatsiooni-restriktsiooni süsteemid bakterites:
- Reparatsioon
- DNA reparatsiooni mehhanismid
- DNA reparatsiooni süsteemid: (tüüp-viga-ensüümid)
- NVR reaktsioonid – Uvr teema
- RNA reparatsioon
- Valkude parandamine
- Homoloogiline rekombinatsioon:
- Transposoonid
- T-DNA: bakterist üle kantav DNA
- Transkriptsioon
- RNA polümeraas
- Promootor
- Erinevad geenid
- Prokarüootidel transkriptsiooni regulatsioon
- Transkriptsiooni initsiatsioon, regulatsioon
- Kromatiini struktuur
- Histoonide modifikatsioonide kaks toimet
- Histooni H3 kovalentsed modifikatsioonid omavad erinevat tähendust
- Eukarüootide pre-mRNA splaissimine - kokkulõikus
- Intron-ekson skeem
- RNA splaissimine
- ATAC intronid ehk minoorsed intronid
- Kotranskriptsioonilised sündmused pre-mRNA sünteesil
- Valgu biosüntees
- Universaalne geneetiline kood
- Lugemisraam ORF (avatud lugemisraam)
- Koodonite kallutatus
- CAI – koodonite adaptatsiooni indeks
- Standardiseerimine
- Koodonite kontekst
- DNA ja RNA koostis
- tRNA sekundaarstruktuur
- tRNA identsus
- Translatsioon
- Initsiatsioon prokarüootides
- Initsiatsioon eukarüootides
- Elongatsioon
- Terminatsioon
- Ribosoomi struktuur
- Ribosoomi komponendid
- Antibiootikumid ja valgusüntees
- Antibiootikumide resistentsuse tagavad erinevad teed
- Mutatsioonid ribosoomides tagavad resistentsuse ABde suhtes
- tmRNA
- Bakterite ja eukarüootide translatsiooni võrdlus
- RNA kvaliteedi kontroll päristuumsetes
- Nonsense mediated decay
- Translatsiooniline kontroll
- No-go decay
- Transkriptsiooni aktivaator GCN4 norm rakus
- Ko-translatsiooniline transport
- Valgu voltumine
- Chaperon
- Hsp 60 suunatud voltumine
- Hsp 70
- Valkude lagundamine
- Proteasoom
- N-otsa reegel
- Hiire fibroplastide kultuur
- Globaalne geeniekspressiooni regulatsioon
- Epigeneetiline pärilikkus ja geeni vaigistamine
- Post-transkriptsiooniline geeni vaigistamine
- miRNA (microRNA)
- miRNAde biogenees
- P-kehad
- RNA interferents (RNAi)
- miRNAd võivad indutseerida mRNA lagundamist
- RNA interferents (RNAi)
- siRNA vaigistamise mehanism
- RISC kompleks
- RNAi kasutamise meetod.
- Transgeensed taimed
- RNAa
- Muul ja Hinni
- Mesilased
- X kromosoomi vaigistamine Xist RNA abil
- CRISPR/Cas süsteem, geenide vaigistamine bakterites
- DNA vermimine isa ja emaliinis
- Prion
- Geenid ja evolutsioon
- Mudel-liigid
- Geeni puu
- Inimlikkuse geen



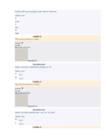









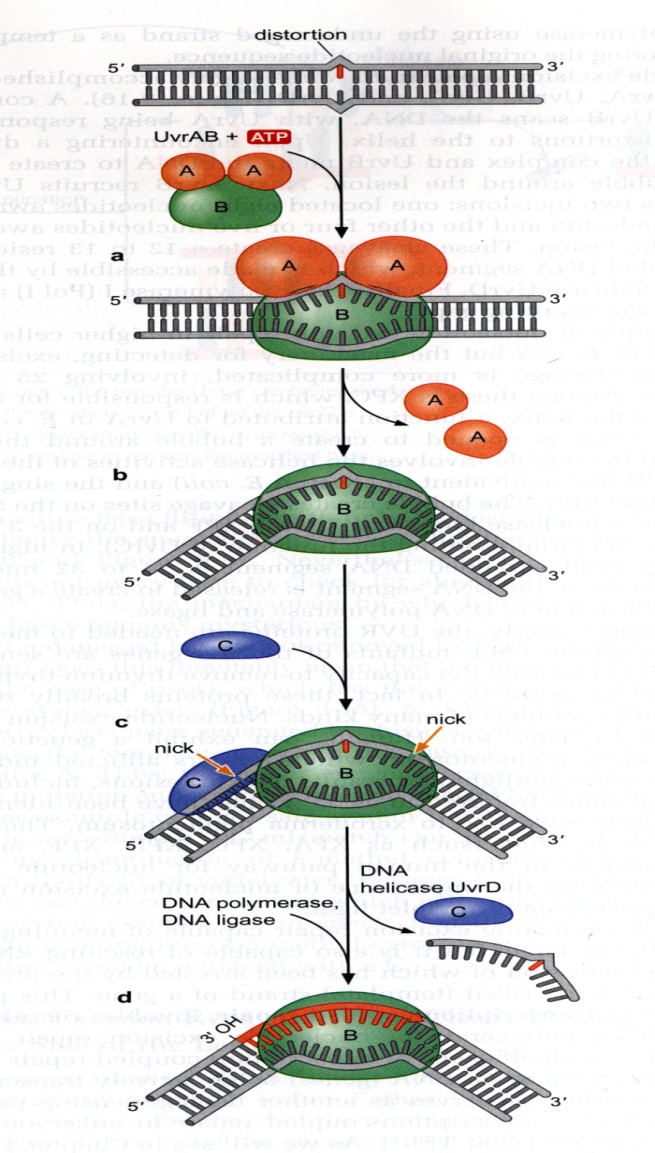
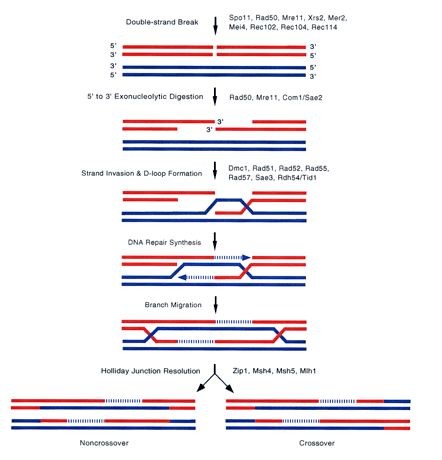


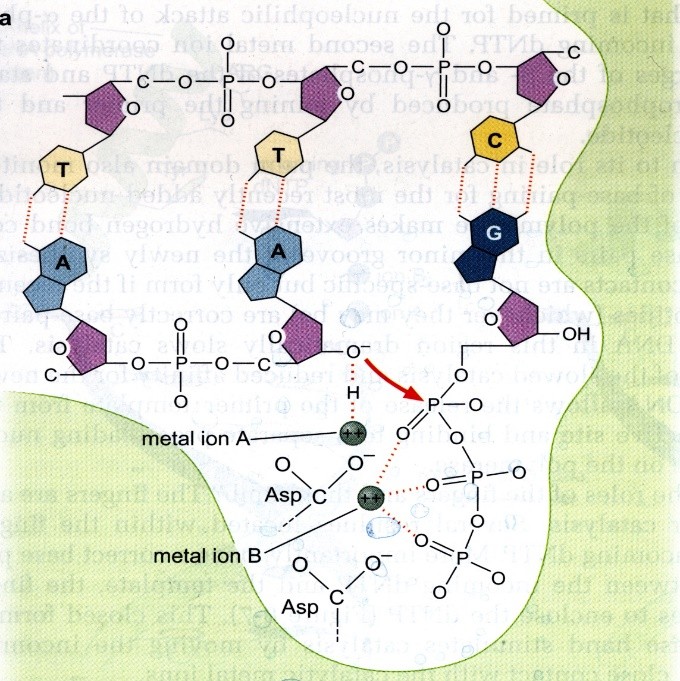
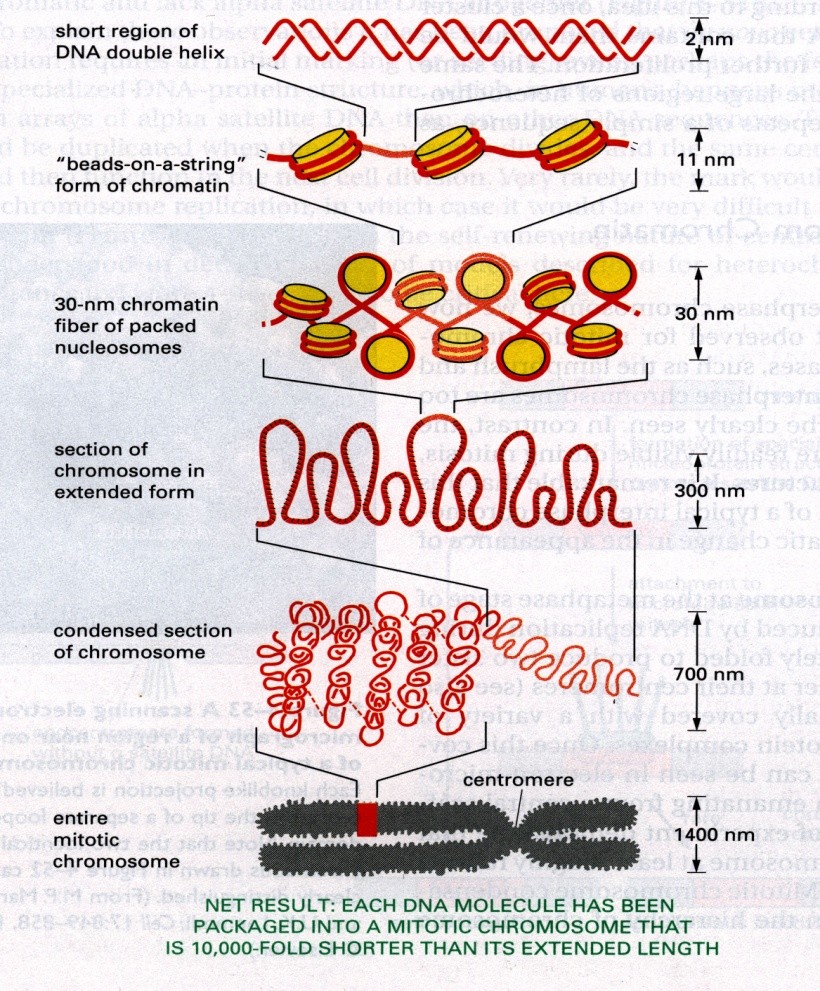
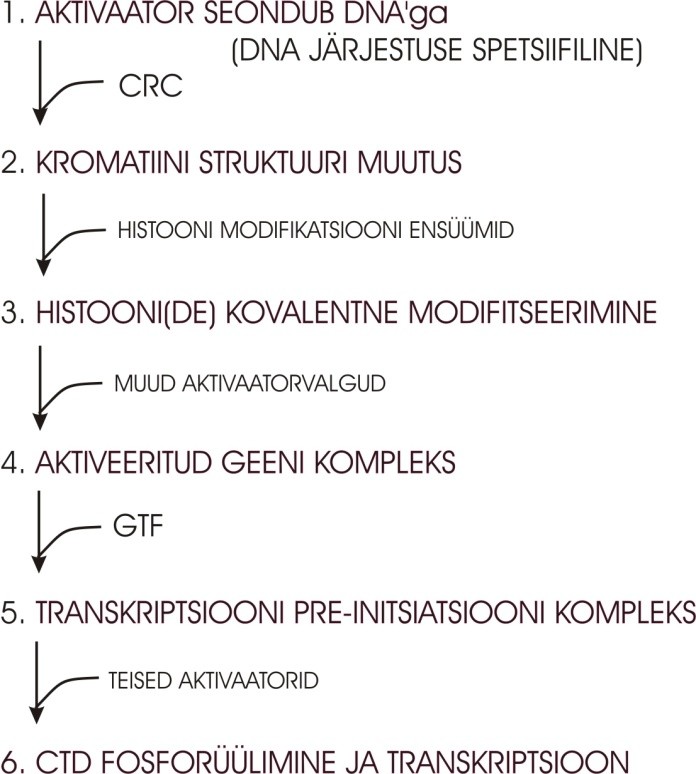
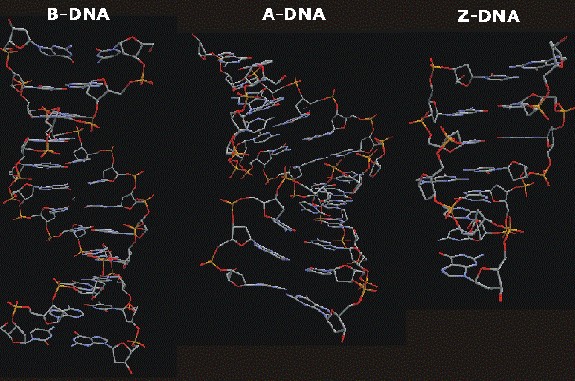


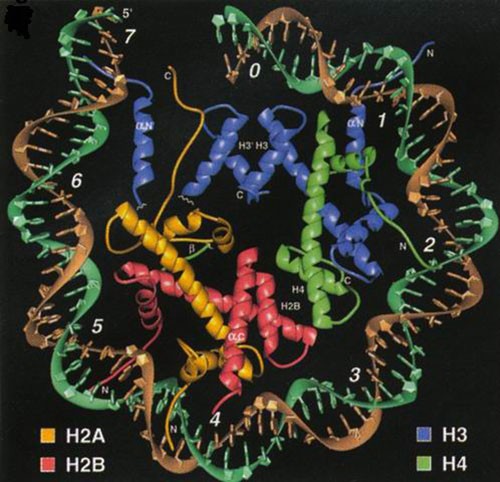
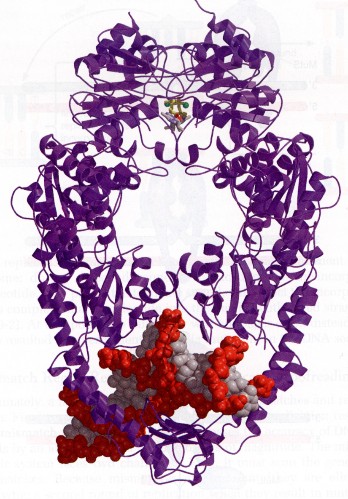
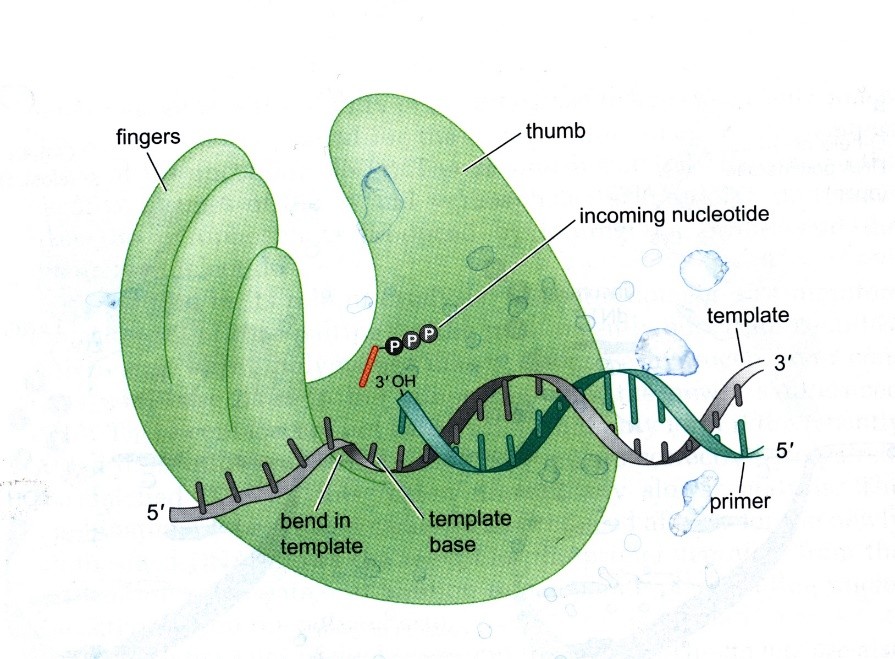
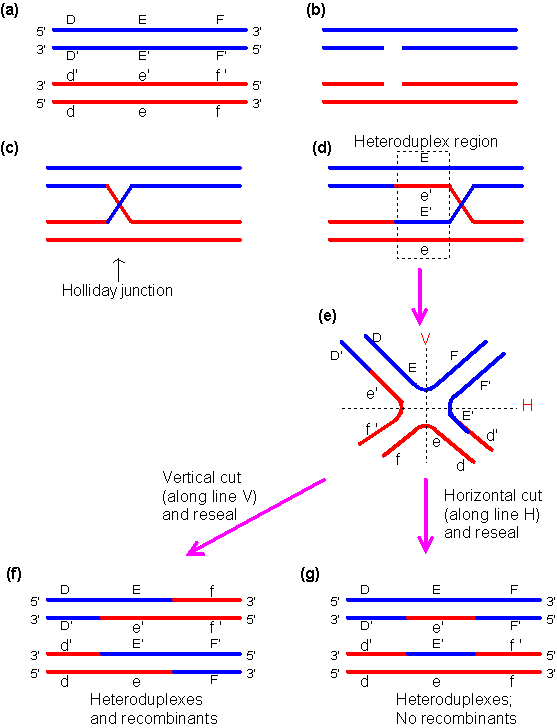
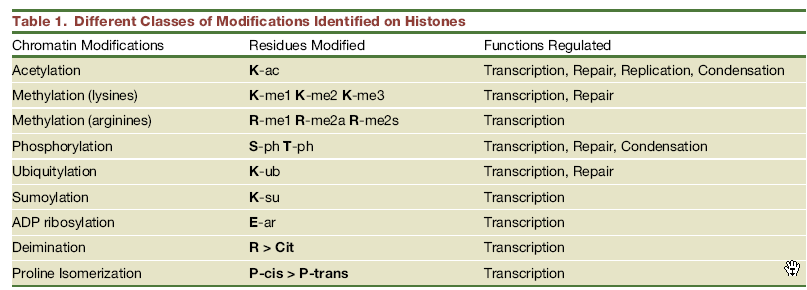















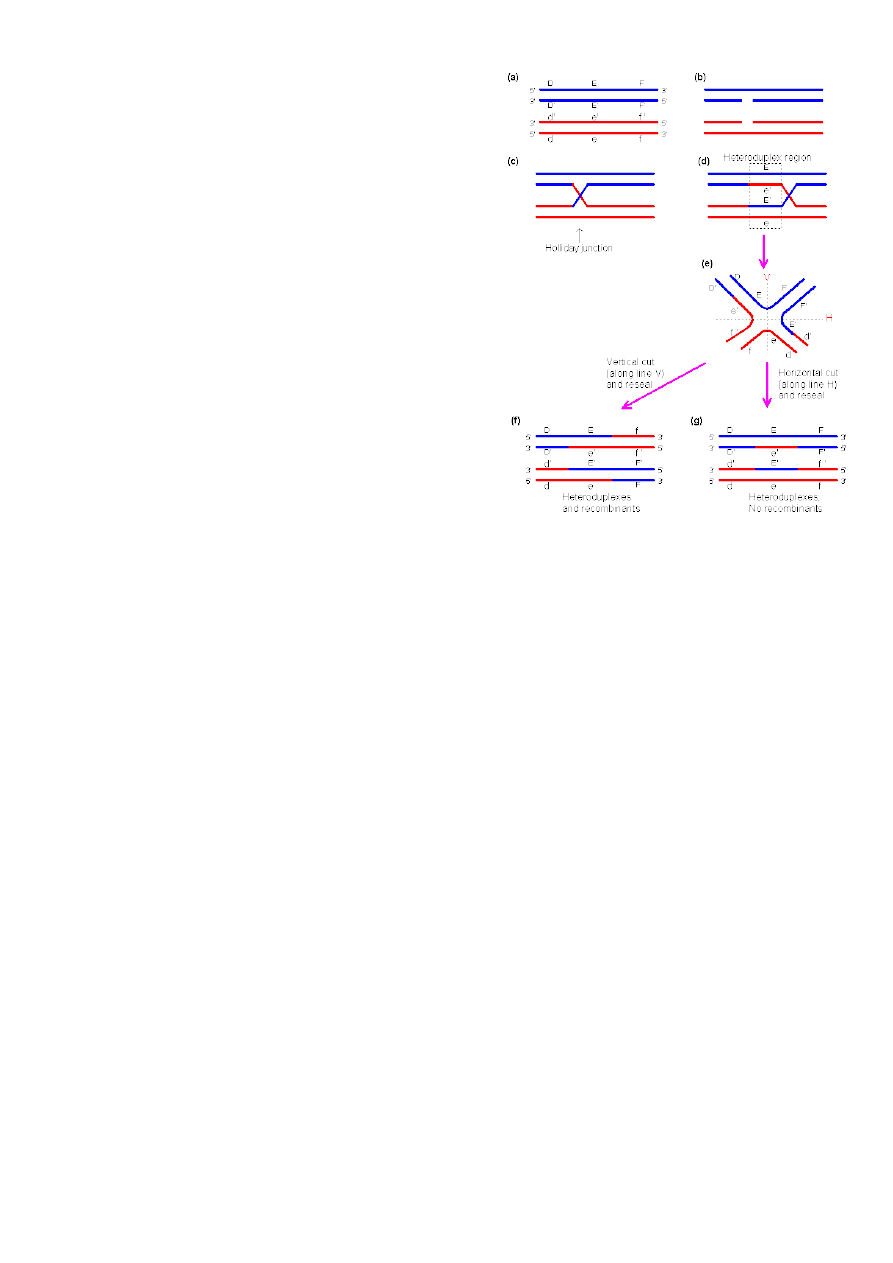
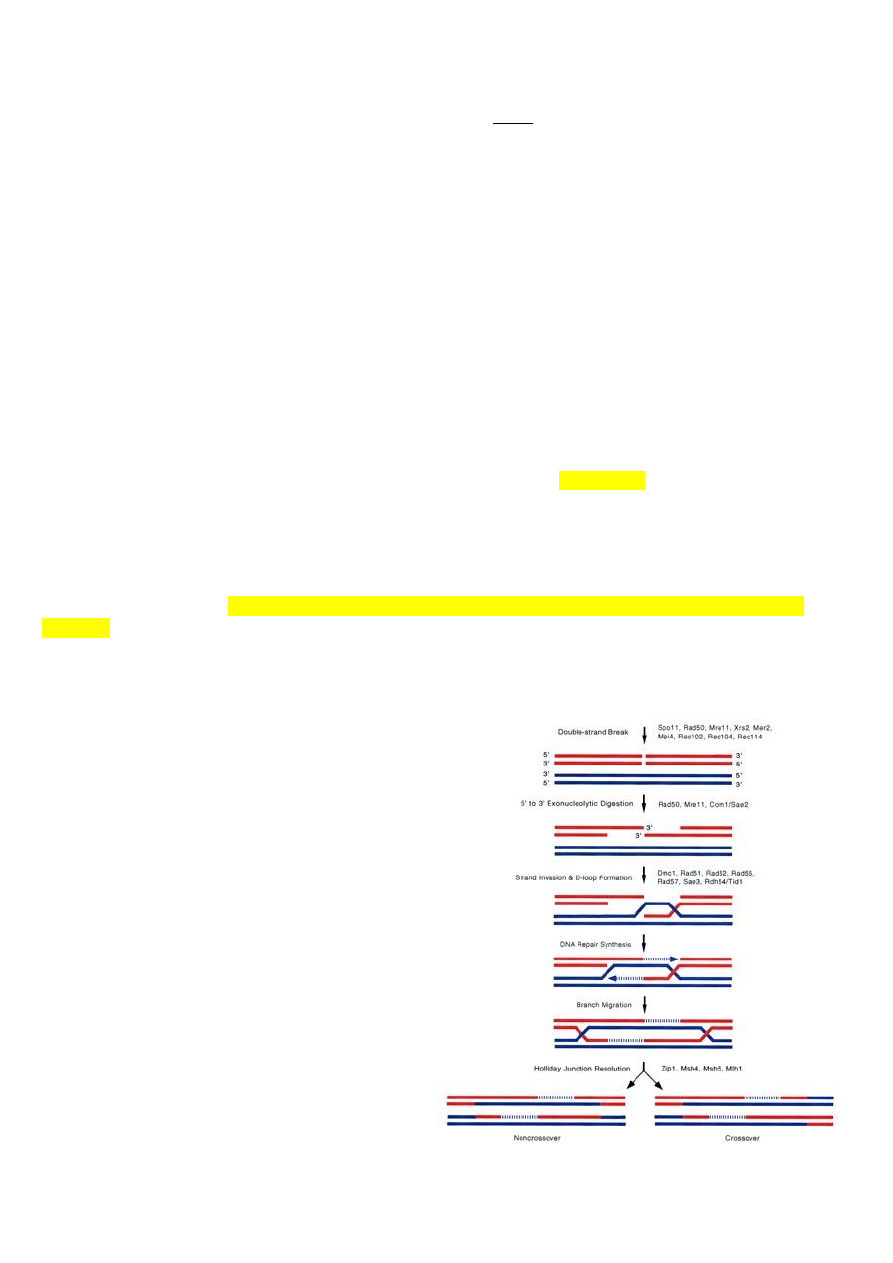




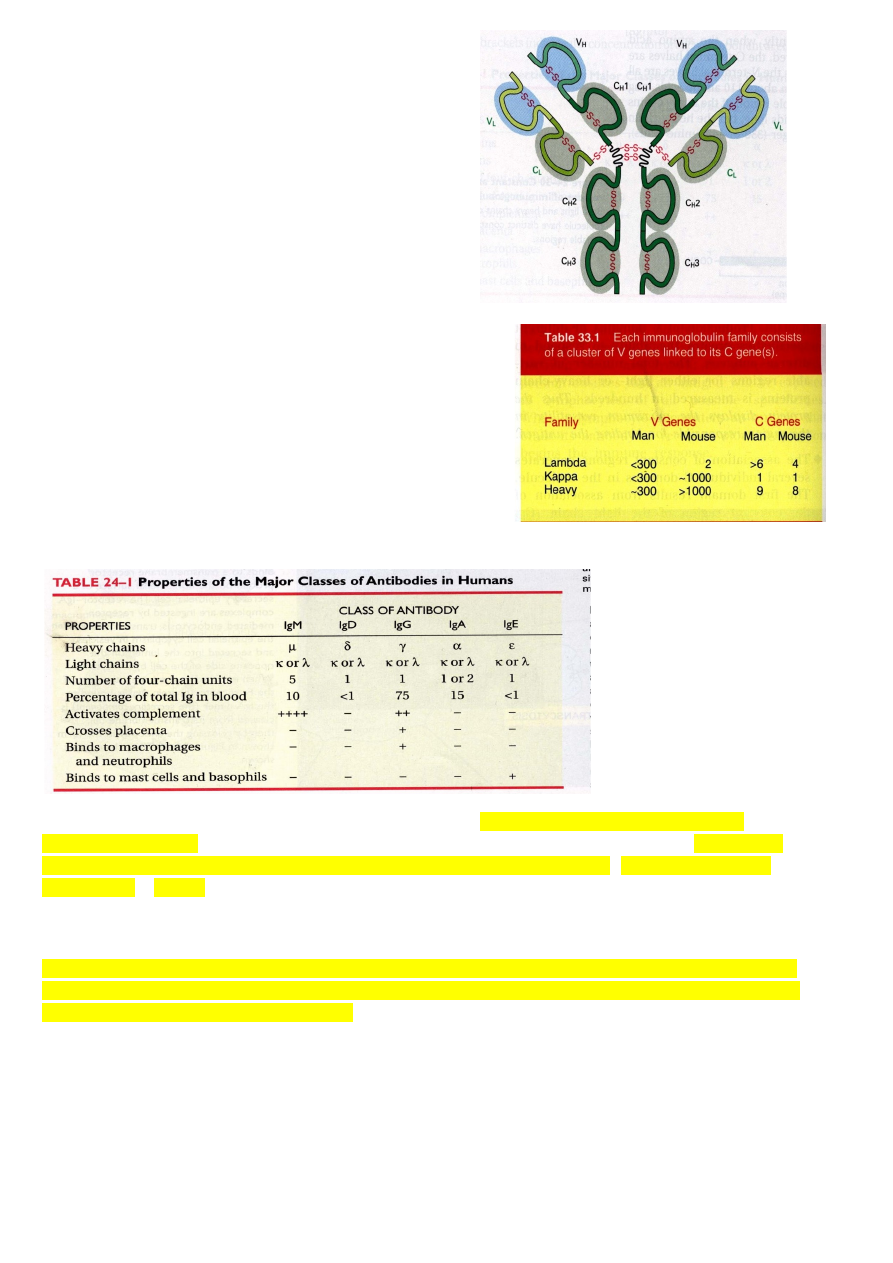







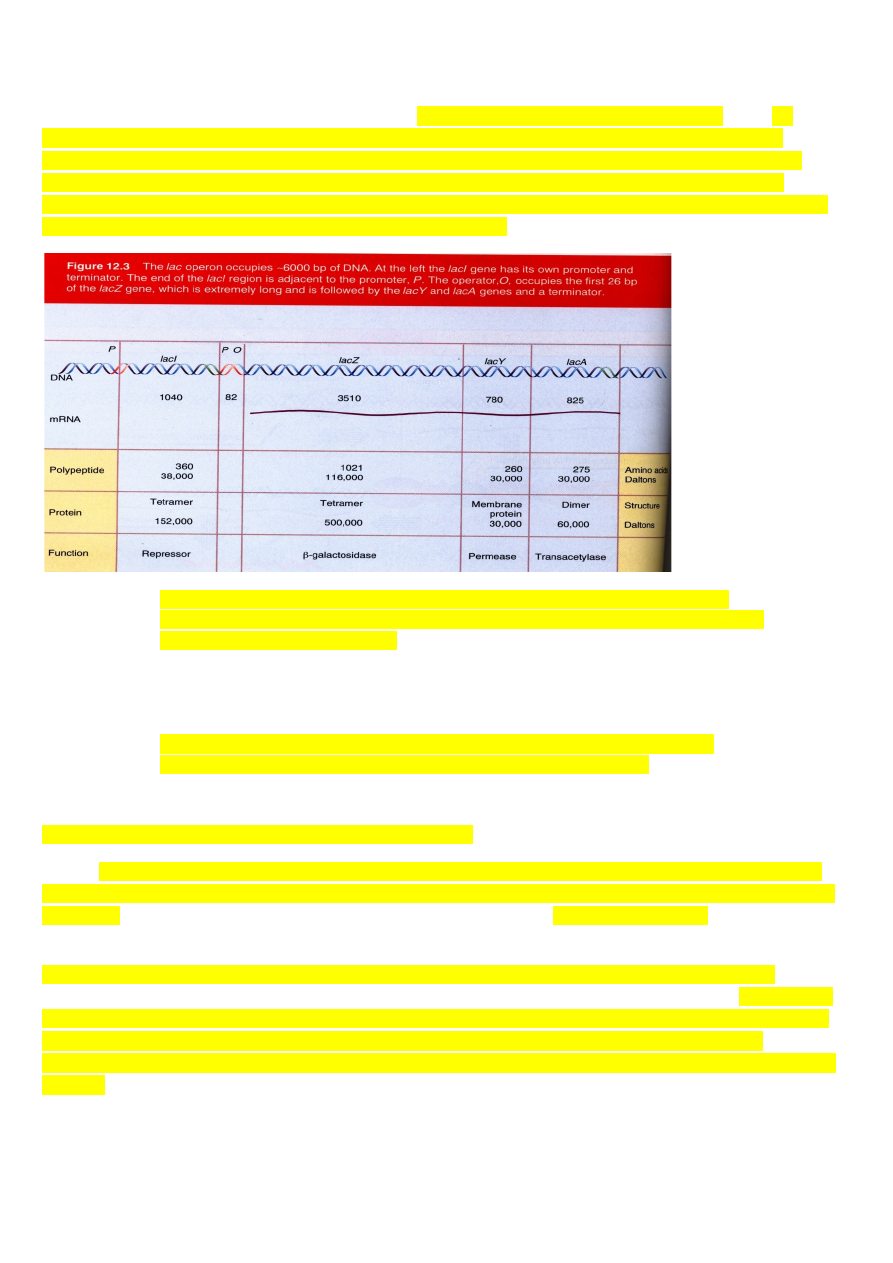










































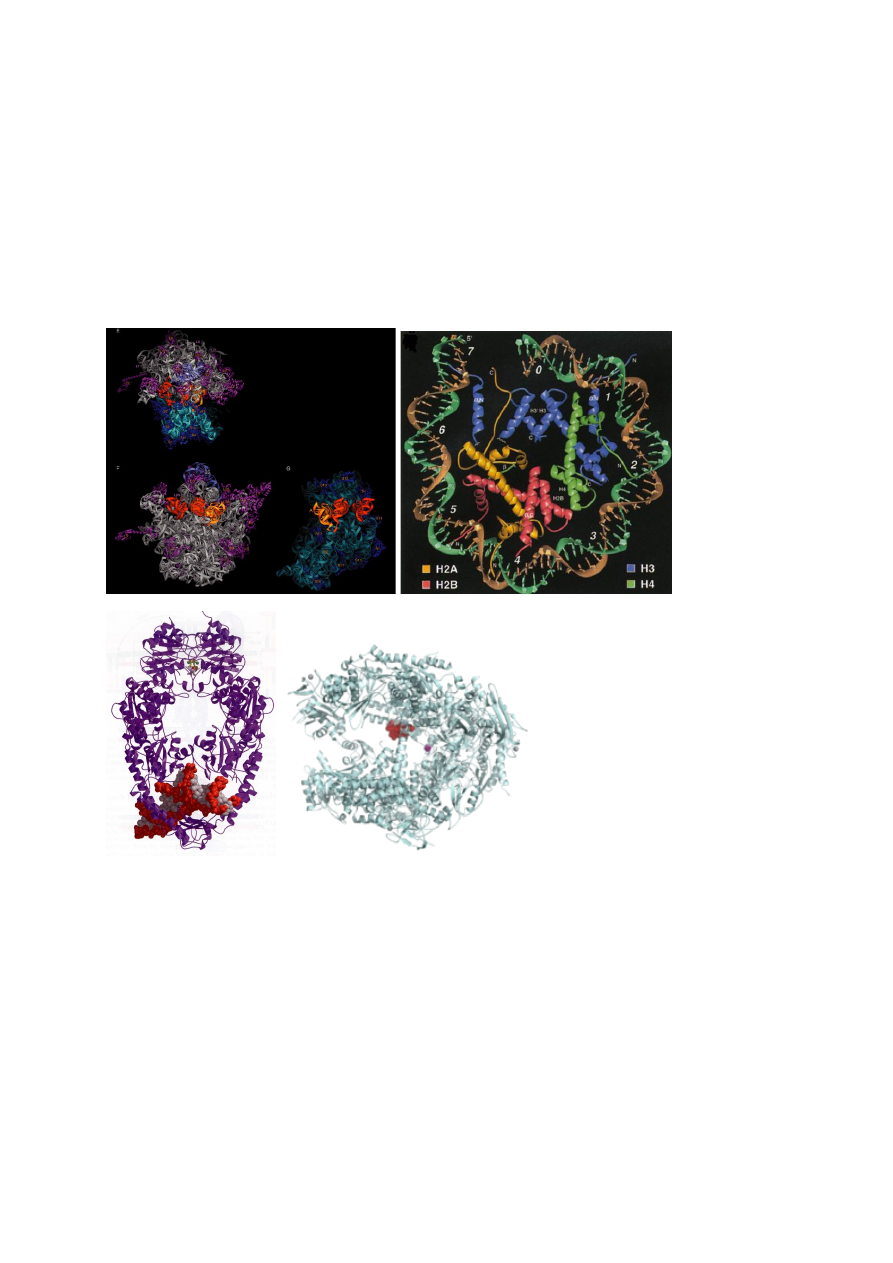




Kõik kommentaarid