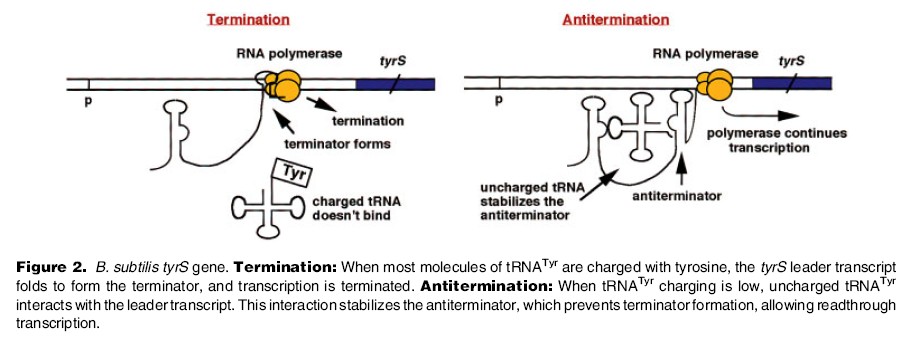
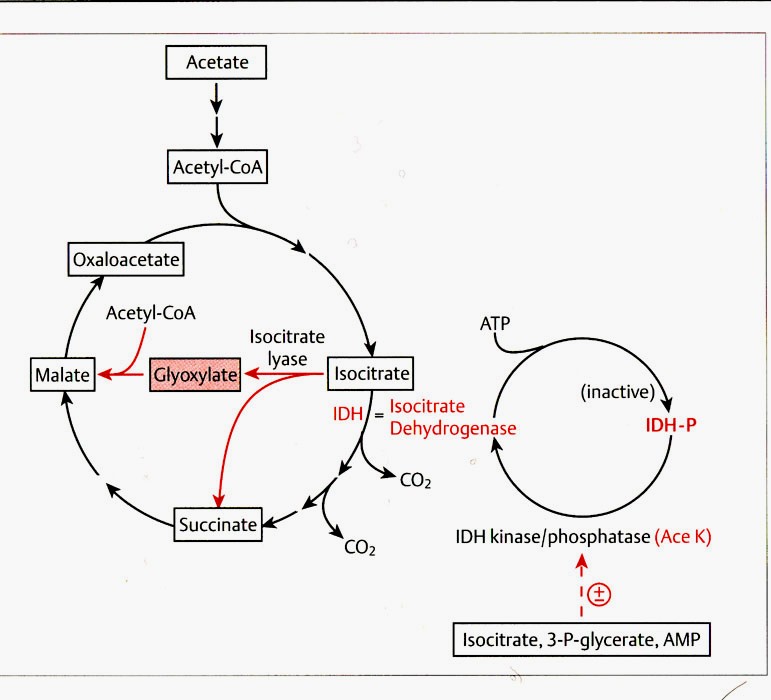
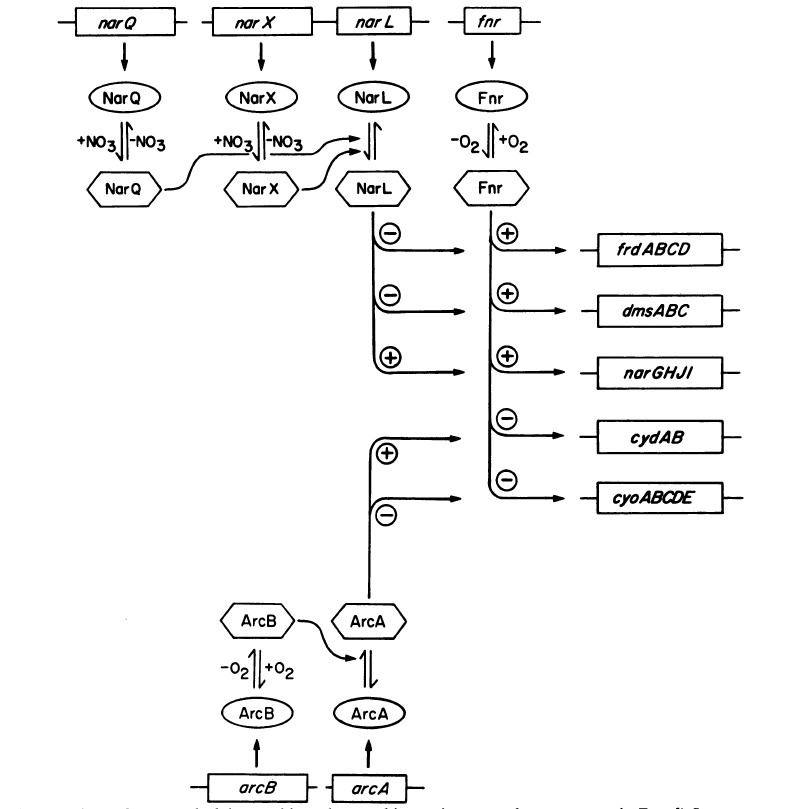
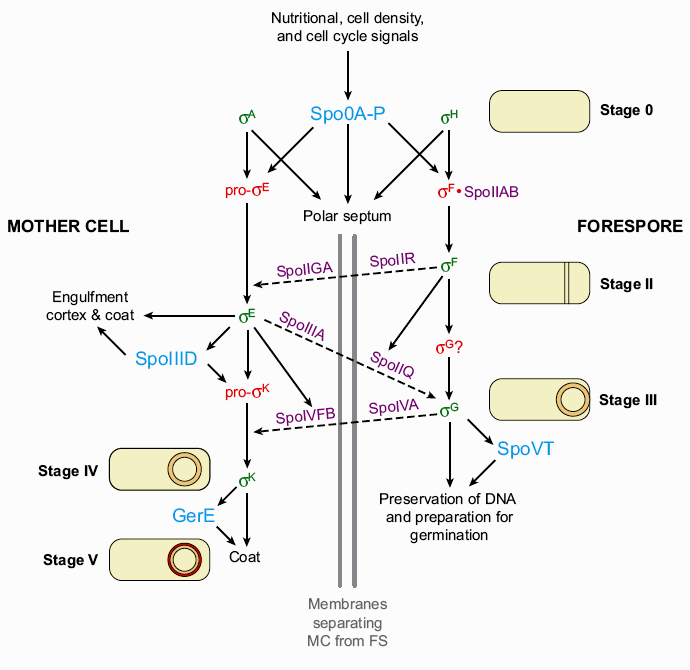
1. Sissejuhatus Metaboolne ja geneetiline regulatsioon bakterites Bakterirakkude efektiivseks kasvuks on vaja, et kõiki raku põhilisi ehitusblokke ja nendeks vajalikke makromolekule produtseeritaks õiges vahekorras. Selleks, et sünteesi lõpp-produktide kontsentratsioon rakus liiga kõrgele ei tõuseks, on rakus välja kujunenud kaks kontrollmehhanismi: 1. Ensüümiaktiivsuse tagasisidestuslik inhibitsioon (feedback inhibition) metaboolne regulatsioon 2. Ensüümi sünteesi repressioon geneetiline regulatsioon Tagasisidestusliku inhibitsiooni tulemusena inhibeeritakse rakus juba olemasoleva ensüümi aktiivsus reaktsiooni lõpp-produkti poolt. Inhibitsiooni võib esile kutsuda ka teatav metabolismiraja vaheprodukt. Geneetilise repressiooni korral inhibeerib tavaliselt lõpp-produkt metabolismiraja esimese ensüümi sünteesi vastava geeni avaldumise pärssimise kaudu. Metaboolne regulatsioon tagasisidestusliku inhibitsiooni kaudu ja geneetiline regulatsioon ensüümi s
4Mikroobifüsioloogia LOMR.03.022 Riho Teras Sisukord 1. Bakterite kasv ja toitumine................................................................................ 4 1.1. Bakterite kasvatamine laboritingimustes.....................................................4 1.2. Elutegevuseks vajalikud elemendid.............................................................7 1.3. Söötmed bakterite kasvatamiseks laboris....................................................9 1.4. Füüsikalis-keemilised tegurid, mis mõjutavad bakterite kasvu...................10 2. Bakterite ehitus ja rakustruktuuride funktisoonid.............................................15 2.1. Tsütoplasma komponendid.........................................................................16 2.1.1. Nukleoid............................................................................................... 16 2.1.2. Tsütoplasma ja inklusioonkehad...........................................................19
viimane veel eriti kondenseerunud vormiks ning lõpptulemus on metastaasi kromosoom. Kromatiini pakkimine: nukleosoomid koosnevad valgulisest tüvest, millele DNA on keermeliselt ümber keeratud, oktameerne tüvi koosneb iga histooni (H2A, H2B, H3 ja H4) kahest koopiast, 30-nm fiibrid, neis on nukleosoomid pakitud irregulaarse struktuurina või solenoidi struktuuri, H1, viies histoon, on solenoidi sisemuses otseses kontaktis DNAga, nii et iga H1 molekul on assotsieeritud ühe nukleosoomiga. Eu- ja heterokromatiin. Eukromatiin on ala, kus kromatiin on vähem kondenseerunud, annab värvimisel heledaid vööte, suurem osa transkriptsioonist toimub eukromatiini piirkondades. Heterokromatiin on ala, kus kromatiin on rohkem kondenseerunud, annab värvimisel tumedaid vööte, esineb sagedamini tsentromeeride piirkonnas ja telomeeride aladel. Kromatiid. Iga metafaasi kromosoom koosneb kahest tsentromeerile kinnitunud tütarkromatiidist. 9
Valgusünteesi etapid eukarüoodi rakus. Transkriptsiooni algatamine, Cap-struktuuri lisamine,mRNA ahela pikendamine, splaissing, mRNA lõikamine, polüadenüleerimine, mRNA sünteesi lõpetamine, mRNA tuumast välja eksport, mRNA lagundamine, Valgusünteesi algatamine, Valgusünteesi lõpetamine, valkude kokkupakkimine, Valkude lagundamine Signaalide ülekanne rakkudes. Rakku saabuvate ja rakus levivate signaalide integreeritus. Intratsellulaarne retseptro molekul kinnitub retseptor valgule, edasi toimub signaali edasikanne intratsellulaarsete signaalvalkude kaudu, kuni signaal jõuab märklaud valguni ning toimub muutus metabolisimis, geeni ekspressioonis või raku kuju muutus ning liikumine Rakkude reageerimine väljast tulevatele signaalidele. Rakk jääb ellu,jaguneb, differentseerub või sureb Rakkudevahelise signaali ülekande viisid. Endokriinne signaliseerimine (hormooni sekreteerimine verre)Parakriinne signaliseerimine-
introni 5' otsa lahtilõikamine, mille tagajärjel introni ees olev ekson lahutatakse ülejäänud mRNA järjestusest. Introni 5' ots keeratakse aasana tagasi ja ühendatakse introni sees olevasse kindlasse järjestusse nukleotiidiga A, mis asub 18-38 nukleotiidi introni algusest 3' suunas. 2 etap introni 3' otsa lahtilõikamine, mille tulemusena intron vabaneb pre mRNA koosseisust. 3 etap allesjäänud eksonite alad ühendatakse ligaaside abil ning tekib ainult eksonitest koosnev mRNA molekul. Intronid degradeeritakse. 36. U snRNAd osalevad pre-mRNA splaisingu regulatsioonis: 5 snRNPd: U1, U2, U4, U5, U6. 37. GT-AT reegel on selline vaatlus, et kõik intronid DNAs algavad GT nukleotiitudega (guaniin, tümiin) ja lõppevad AG nukleotiitidega (adeniin, guaniin). Kui DNA on transkribeeritud RNAsse, intronid on eemaldatud RNAst mehaniismi abil, mis tunneb ära neid nukleotiitide alguseid ja lõppe RNAs nad oleksid CU (tsütosiin, uratsiil) ja AC (adeniin, tsütosiin). 39
1 MOLEKULAARBIOLOOGIA. 1. Kui aatom loovutab elektroni täielikult teisele aatomile, missugused keemilise sidemega on tegemist? Ioonside, sellised ained lahustuvad hästi, kuna ioonide hüdratatsioonienergia on suurem kui kristalli võreenergia 2. Miks vesi on hea lahusti (solvent)? Vesi on hea lahusti, sest ta lahustab nii tahkeid, vedelaid kui ka gaasilisi aineid. Vee molekul moodustab dipooli ning aatomid omandavad osalise laengu. Polaarsete ühenditega moodustab vesiniksidemeid, mis tagavad stabiilsust. 3. Termodünaamika II seadus. Kõik protsessid kulgevad tasakaalu e. minimaalse potentsiaalse energia poole e. entroopia kasvu suunas. Entroopia (S) on korrastamatuse mõõt [J/mol*K], korrastatud madal entroopia. Isoleeritud süsteemid püüavad korrastatud olekust korrastamata poole. Tasakaal on siis, kui entroopia on maksimaalne.Entroopia muutus on
2) RNA protsessimine ja transport. Aktiivne protsess ja väga täpselt kontrollitud 3) mRNA lagundamine - mRNA eluiga on oluline. Kui seda ei lagundata, siis saab tema arvelt sünteesida palju komponente. Valk titiin (kõikidel selgroogsetel olemas) – vajatakse varajases embrüonaalses arengus, esimest korda vajatakse blastulas. Sünkroonjagunemine toimub kiiremini (valk valgus sünteesitakse), mRNA jõutakse valmis sünteesida ja rakku transportida, aga ei jõuta lõpuni transleerida. Titiini molekul ise on väga pikk (mitukümmend AH pikk) - jääb titiin poolikuks, sest mRNA 3 lagundatakse enne mitoosi käigus. Kui rakutsükkel pikeneb, saavad titiini molekulid valmis. 4) translatsioon – mRNA struktuur ja regulaatorvalgud ja RNA-d 5) valgu modifitseerimine ja lokalisatsioon – erinevatel modifitseerivatel valkudel eri roll 6) valgu eluiga (N-terminaalne reegel) – valke sünteesitakse tohutult palju, aga kui
(viiruse kapsiidi struktuurvalk) ja pol (pöörtranskriptaas/integraas) geenidele. Ty1 element on leitud pagaripärmist (saccharomyces cerevisiae); Ty element on 5,9 kb pikkune, sisaldab 340 bp pikkuseid LTR-e. Tekitab transpositsioonil 5 bp pikkuseid otsekordusjärjestusi. Pärmi genoomis on leitud 35 koopiat Ty elementi. TyA ja TyB (pöördtranskriptaas ehk revertaas, mis sünteesib Ty elemendi RNA-lt dsDNA) on gag ja pol geenide homoloogid. DNA molekul inserteerub genoomi, tekitades sel viisil Ty1 elemendi koopia genoomi teises piirkonnas. Ty moodustab viiruselaadseid partikleid. Retroposoonid: Äädikakärbses on kirjeldatud F, G ja I retrosposonid. Imetajatel on levinud LINE-d (long inserted nuclear elements, 6-7 kb). Retroposoonide otstes pole LTR-e, kuid nende ühes otsas on homogeenne A:T aluspaaridest koosnev järjestus. A:T järjestus liitub retroposooni
































































Kõik kommentaarid